Just Another Year in the Genetosphere
The War of Conservatism and Liberalism
by Charles Brack
The grinding stress of population genetics
on political borders:
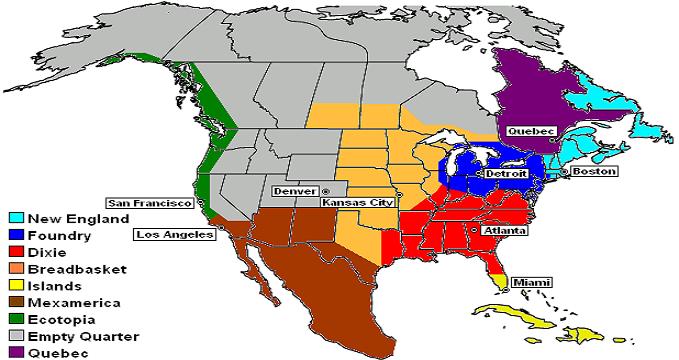
Joel Garreau's Nine Nations of North America
It never ends. Conservatives and liberals had another difficult year, and the cooling down period that usually occurs after a presidential election didn't last very long. The war over health care accelerated during the hot summer months, with a bill finally passing in the Senate three days after the winter solstice.
This is an interesting coincidence, given the proposed relationship between sunlight exposure and political behavior (see Cycles of Conservatism and Liberalism). Briefly, this theory proposes that increased sunlight exposure and heat promote more conservative attitudes and behaviors. Does liberal legislation have a better chance of passage during the winter, following the general increase in social altruism that is punctuated by the Christmas holidays?
The battle over health care highlights the fundamental Darwinian difference between conservatives and liberals: liberals organize their prosocial behavior over greater genetic distance than conservatives. Both conservatism and liberalism induce changes in population gene frequencies. The health care debate highlights this interesting interaction of politics, religion, and genetics, which we shall explore more closely in the upcoming year. But for now, let's look back at some of the more notable theories presented in Neuropolitics.org in 2009.
The thermodynamics of conservatives and liberals
As should probably be expected by the conservative-liberal dichotomy of environmental attitudes, conservatives run thermodynamically hotter, that is, they tend to use more energy. This is fundamentally a reflection of the increased rate of conservative reproduction and their increased demands for space. We estimate that the average conservative produces 40-55% more children than the average liberal. However, this statistic only holds for caucasians.
The thermodynamic divergence of conservatism and liberalism is seen in other species. To quote ourselves: "species that can modulate their thermodynamic properties in response to environmental changes dramatically increase their probability of survival. In humans, there is no better example of thermodynamic modulation than conservatism and liberalism."
The conservative contribution to human species survival is the tendency to increase human habitat range and provide for effective reproduction in smaller social groups, mirroring the phenomenon of r-selection as described by MacArthur and Wilson in their landmark book, The Theory of Island Biogeography (1967). The liberal contribution to species survival lies in the propensity to increase habitat life and support higher equilibrium levels of human biomass in those habitats.
Liberalism and conservatism are simply the human manifestation of r-K selection as originally developed by MacArthur and Wilson to apply to animal and plant species (see The Biogeography of Political-Religious Affiliation). But things are a bit more complicated in the human model, in that the modern human implementation of r and K strategy is symbiotic.
 |
|
This human symbiosis of r and K strategy is executed by the division of labor and the development of habitat corridors linking urban, suburban, and rural areas. In the field of population ecology, a habitat corridor is simply an easy mode of travel between two connected habitats, which increases both the number of species and species viability in both habitats.
In humans, habitat corridors are roads, freeways, subways, canals, rivers, lakes, oceans, air routes, etc., which provide for the rapid transportation of economic resources that mutually support the diversity of the human habitats they connect. Thus, the r strategy of conservatism and the K strategy of liberalism can both be facilitated by habitat corridors between suburban, rural, and urban areas.
The asymmetric geographical distribution of conservatives and liberals seems to be a more effective model of total reproduction than a symmetric distribution. The urban tendencies of liberalism and the suburban-rural tendencies of conservatism provide a degree of isolation of their diverse reproductive strategies, thereby reducing mutual interference. The asymmetric geographical distribution of conservatives and liberals point directly to the r-K dichotomy of conservatism and liberalism.
The political mystery of the DRD4-7 allele
One of the genes frequently implicated in political-religious disposition is the highly polymorphic dopamine receptor gene, DRD4. DRD4 is one of the most polymorphic genes in the human genome, and interestingly, one of the most behaviorally variant. The dopamine system contains a number of different receptor types, and the D4 receptor is most likely to line the neural pathways associated with the politically and religiously hot dopamine reward system.
The
7-repeat allele (DRD4-7) of this gene has been implicated with
migratory behavior (Chen, 1999). Further, long alleles of DRD4
have been implicated, although equivocally, in novelty-seeking,
exploration, early initiation of first sexual activity, desire
for children, drug-dependence, reward-seeking, extraversion,
hyperactivity, spiritual transcendence, OCD, ADHD, and schizophrenia
(in caucasians).
The 7-repeat allele seems to have an inhibiting impact on the experience of reward, inducing greater risk-taking to stimulate the same chemical impact as the more efficient 4-repeat version. Migratory behavior is often risky, and possibly facilitated by higher levels of the 7-repeat. However, this gene has also been implicated in liberalism by James Fowler et al. (see Friendships Moderate an Association Between a Dopamine Gene Variant and Political Ideology), via a proposed environmental interaction with "teenage friendships".
Robert Haston, the author of The Origin of Political Species, has proposed that the DRD4-7 allele actually can contribute to either conservatism or liberalism based on its diverse interactions with variations in population density.
Are political borders performing Darwinian functions?
The origin of the political borders that divide the world's 194 sovereign nations seem to be somewhat arbitrary, but there is a Darwinian method to this madness. Political borders, much like habitat barriers, inhibit gene flow, isolating populations and inducing genetic drift on opposite sides of the border. This interesting phenomenon was noted in a study by Kayser et al. (2005), which found substantial Y-chromosome variations between German and Polish populations following the general rule: the genetic border between Germany and Poland matched the political border quite well. Further, the political border between Germany and Poland is built along the rivers (i.e., natural habitat barriers) that separate the two countries.
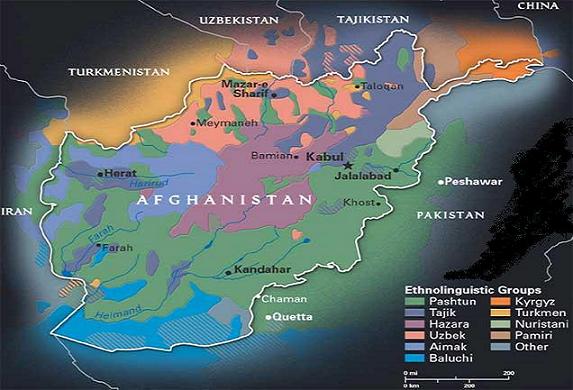
The tribal map of Afghanistan: are political borders
unstable if the genetics on opposite sides of the border are
very similar? The Afghanistan-Pakistan border, which bisects
the Pashtun ethnic group, allows for easy migration.
Political borders are often based on natural habitat barriers, such as mountains, rivers, lakes, and oceans. This exposes the Darwinian implications of political boundaries, as they restrict gene flow, resulting in genetic drift between populations on opposite sides of the border. Political borders function as habitat barriers, and the gene flow across a political border varies in proportion to the ease of migration across that border.
Therefore, countries that severely restrict immigration, such as North Korea, are especially susceptible to genetic drift. The proposal that political borders function, in part, to preserve population gene pools certainly has merit.
Capitalism, communism, and population genetics
One of the overlooked characteristics of economic systems is their impact on population genetics. Capitalism is the Darwinian buzz saw of economic systems, inducing rapid shifts in population gene frequencies, especially as it is introduced to new populations (see The Evolutionary Pressure of Profit). One of the ways capitalism generates profits is by reducing the energy utilization differences between populations. This is best seen in the continual movement of manufacturing to low-wage populations, and the growing plight of the American worker.
Interestingly, communism is best executed in social groups with low genetic distance, and functions quite well within the confines of the nuclear family. Indeed, most ardent anti-communists unknowingly practice communism every day of their lives, often known as the "nuclear family". These anti-communists only notice communism if it is combined with genetic distance, and the application of selfish-gene theory to political behavior is quite appropriate in their case.
As genetic distance grows, communism becomes problematical to execute. Communism therefore benefits by restrictions to migration and father-like leaders that maintain power for long periods of time.
Skin color and political disposition
Of all the theories we introduced in 2009, the proposed biological relationship between skin color and political disposition drew the most fire (see Latitude). The theory is founded upon the rather interesting fact that both the brain and skin evolve from the same embryonic structures. Thus their unusually close relationship, which makes sense, given the skin's supporting role in the detection of temperature, pressure, touch, vibration, and pain. This is also seen in the close relationship between stress and skin disorders such as shingles, herpes, and psoriasis.
This runs right into the monoamine hypothesis of conservatism and liberalism, where the serotonergic and noradrenergic systems are oriented towards behaviors and attitudes generally categorizable as liberal, while the dopaminergic system is more oriented towards conservatism.
Curiously, cutaneous melanocytes, which produce melanin in the skin, also produce serotonin, which facilitates the darkening of the skin. Further, serotonin receptors are expressed in melanocyte cells, an indication that serotonin is used in the bioregulation of skin color. However, the linkage between skin color and liberalism is reliant on the proposed relationship between skin, brain, and blood platelet serotonin. In humans, platelet serotonin uptake site and brain serotonin transporter are coded from the same single-copy gene, located on chromosome 17 (Lesch, 2006).
Further, dopamine, which we have proposed to be more oriented towards conservative behaviors and attitudes, has been found to inhibit the expression of melanin in the skin. Thus, dopamine and serotonin seem to have opposite impacts on both skin color and political disposition. Genes impacting melanin physiology have been under considerable selective pressure, and this might have spilled over into cognition.
Noradrenaline, another neurotransmitter that we suspect contributes more towards liberal attitudes and behaviors, has also been linked to the inhibition of the expression of melanin in the skin. This certainly presents a dilemma to the theory, except for the fact that it might explain some of the distinctive behaviors and attitudes of caucasian liberalism, in particular, its lower tolerance for population density and its extraordinary sensitivity to environmental contamination. More on the diverse influences of the serotonergic and noradrenergic systems on political-religious behavior upcoming this year in Neuropolitics.org.
Previc's general theory of neurotheology
One of the more interesting and pervasive characteristics of modern religions is their orientation towards the upper visual field. The positioning of god and heaven into upper, distant space is not so much about god and heaven as it is about the way the brain organizes space (see God, Dopamine, and 3-Dimensional Space).
 |
Why are God and Heaven positioned
in the upper visual field? Religions are ubiquitous
in their positioning of the good gods and religious objects
into upper space. This riddle was resolved by Fred Previc,
who proposed that neural networks, predominately in the
ventral dopaminergic pathways in the left hemisphere,
are predisposed to the processing of distant, upper space.
These same neural networks have been closely linked to
religious behavior in many studies, thus the curious relationship
between religion and the upper visual field. |
While the Persinger and D'Aquili-Newberg models of religious experience were ingenious, the many diverse phenomena associated with religiosity were still confusing and seemingly disconnected: the high rates of psychopathology associated with religious conversions, dreams, hallucinations, and awakenings; the organization of God and heaven into upper, distant space, and the Devil and hell into lower space; the heavy role of incentive-motivational behavior associated with religious activity; the orientation of the religious to seeking rewards in distant space and time (i.e., heaven); the close correlation between religiosity and political conservatism; and, the interesting correlations between religious thought and general abstract reasoning.
Fred Previc, a leading expert in the field of human spatial disorientation during flight, proposed that the brain maps and directs its behaviors in four distinct realms of space (1998), each of which has specific neural networks that simultaneously work to optimize the various categories of behaviors we engage in, and seems to us to be a single 3-dimensional world. From this, and the D'Aquili-Newberg model of religious experience, Previc put it all together. God isn't so much about God as he is about the way the brain works, and the peculiar human version of God echoes this neural organization.
Previc's dopaminergic-spatial theory is the general theory of neurotheology, with the Persinger and D'Aquili-Newberg models becoming special cases. Previc granted us an interview in 2009 (see In the beginning...), and gave this fascinating account of the development of his theory:
The application of my four-dimensional neuropsychological theory of space to religious behavior occurred a few years after its publication in 1998, although I did allude in that paper as to how the primarily dopaminergic ventral areas dealing with extrapersonal space might be involved in even more spatially and temporally distant concepts such as “heaven” and “eternity”. And, I was aware of the religious themes so often reported by paranoid schizophrenics, in concert with the well-documented overactivation of dopaminergic pathways of the left hemisphere in that disorder. In the early part of this decade, some seminal findings of Newberg and others really advanced our understanding of the brain areas involved in religious experience and pointed to a greater ventral prefrontal and temporal-lobe involvement, so I began an extensive review of the neuropsychology (and particularly neurochemistry) of religious behavior. One important aspect of the theory I ultimately presented was the association of “religious experience” systems in the brain to those underlying a variety of extrapersonal states (including dreaming and hallucinations) in which a de-emphasis on tactile and other bodily sensory inputs occurs. Hence, religious systems, like most other brain systems, are not unique in and of themselves but derive from a more fundamental organization of the brain in terms of how we interact in different regions of space. Perhaps this is what most other neuropsychological theorists are less willing to accept. It is the same with my evolutionary theories of human intelligence – rather than focus on the evolution of intelligence or language per se, as many theorists have done, I see more fundamental physiological changes at work that produced a variety of uniquely human traits.
The genetics and energetics of class warfare
Nothing quite says genetic conflict among humans like class war, and indeed, humans exhibit both genetic and phenotypic variations across social "classes". Since defining "class" is arbitrary, in 2009, we introduced the "trait" variation model of social class definition (see Trends in Class Warfare). Using this classification mechanism, we noted a number of genetic trends we believe to be associated with class warfare.
Gene flow between classes varies inversely to the genetic distance between them. That is, the greater the genetic distance between two classes, the slower the rate of gene flow. Further exacerbating the problem of gene flow, the greater the genetic variation in a population, the greater the variation in energy utilization in that population. When energy utilization diverges between two populations, so does gene flow.
It also follows that increased population size increases genetic variation, along with increases in social parasitism. Further, the lower class is on an accelerated life cycle, which counters their energy deficit. Changes in technology can induce changes in gene flow between classes. But most importantly, genetic variation across social classes seems to be following the rules of genic selectionism, that is, selfish-gene theory.
The general theory of political-religious orientation
One of our more interesting proposals of 2009 involves the relationship between political-religious orientation and the brain's rendering of the experience of reward (see Political-Religious Disposition and the Dopamine Reward System). Political-religious disposition may be a state of optimizing the experience of reward in the brain. That is, a person's political-religious opinions optimize the experience of reward in the dopamine system.
There are several studies which support this proposal. The Harbaugh experiment, Neural responses to taxation and voluntary giving reveal motives for charitable donations (2007), found that both voluntary and involuntary charitable contributions (taxation) activated the dopamine reward system, increasing the experience of reward, particularly in the right hemisphere. This provides yet more evidence that the tax-tolerant liberals are more likely to be under the influence of their right hemispheres.
Further, the Schjødt experiment, Rewarding prayers (2008), found that praying also increases the experience of reward in the brain, but only for those that believe strongly in God, and further, that God answers prayers. Interestingly, this activated the caudate nucleus in the right hemisphere (part of the dopamine reward system), which has been specifically linked to the prediction of rewards associated with reciprocal social behavior, that is, rewards associated with others returning favors. This is a common element of prayer, in which one seeks to strike a "deal" with God.
Another study in support of the hypothesis that political-religious opinions optimize the experience of reward in the brain was the Westen experiment (2006). In this experiment, conservatives and liberals were caught in the act of activating their dopamine reward systems in response to positive information about their favorite political candidates. The activation of the dopamine reward system was maximized when negative information about their candidate was presented, followed by exculpatory information.
The benefit of a reward theory of political-religious disposition is the richness of political-religious disposition it can explain. Since the dopamine reward system is one of the major integrative regions of the brain, with reciprocal connections to the prefrontal and insular cortices, striatum, hippocampus, amygdala, PAG, thalamus, hypothalamus, etc., it provides the foundation for the diversity and complexity in political-religious disposition.
There are indeed many flavors of conservatism and liberalism, which might be explained by the relative influences the various regions of the brain weigh upon the dopamine reward system in the right and left hemispheres. More "intellectual" versions of the conservatism and liberalism might be the result of the greater influence of the left and right prefrontal cortices, respectively, on the dopamine reward system.
Just another year in the genetosphere
One of the fundamental hypotheses of political conflict is that it increases with genetic distance. The fundamental unit of natural selection is not the human, which may survive a hundred years, or the genome, which is badly fragmented by the process of meiosis. The fundamental unit is the gene, and we are unknowingly living our lives for them.

The Darwinian moment: waking up, only to realize your life
has been a complete illusion
This wouldn't be so bad, except for their never ending conflict. Gene competition, or genetic drive, occurs at all levels of biological interaction: genes within a genome; cells within an organism; organisms within a population; populations within a species; and, between species. This fundamental genetic conflict is also expressed in political behavior.
Beauty, taste, sex, love, and even god are servants to the selfish genes. A entire illusory world has been constructed for us, which we call the genetosphere. The genetosphere is the neurobiology that has been constructed to support the replication of the selfish genes, and something akin to the illusory world created by the robots in the movie The Matrix. The genes are responsible for the constant state of conflictedness in life, and we are caught in the crossfire of their ugly and never ending wars of attrition.
As we enter the final year of Neuropolitics.org, we will finish our study of the eternal and fundamentally genetic battle of conservatism and liberalism. As we are subjected to the ongoing battle over health care, the emerging battle over cap and trade, and the war over the continuation of the Bush tax cuts--remember, it is just another year in the genetosphere. And, unlike the movie, there is no waking up.
Charles Brack, January 2010.
References:
C. Chen, M. Burton, E. Greenberger, and J. Dmitrieva (1999). Population migration and the variation of dopamine receptor (DRD4) allele frequencies around the globe. Evolution and Human Behavior, 20, 309-324.
W. Hamilton (1963) The evolution of altruistic behavior. The American Naturalist. 97: 354–356.
W. Harbaugh, U. Mayr, and D. Burghart (2007). Neural responses to taxation and voluntary giving reveal motives for charitable donations. Science 316, 1622–1625.
M. Kayser et al. (2005) Significant genetic differentiation between Poland and Germany follows present-day political borders, as revealed by Y-chromosome analysis. Human genetics. 2005, vol. 117, no5, pp. 428-443.
K. Lesch, B. Wolozin, D. Murphy, and P. Riederer (2006) Primary Structure of the Human Platelet Serotonin Uptake Site: Identity with the Brain Serotonin Transporter. Journal of Neurochemistry. Volume 60 Issue 6, Pages 2319 - 2322.
R. MacArthur and E. Wilson (1967) The Theory of Island Biogeography. Princeton, NJ: Princeton University Press.
F. Previc (1998) The neuropsychology of 3-D space. Psychological Bulletin. Psychological Bulletin. 1998 Sep;124(2):123-64.
U. Schjødt, H. Stødkilde-Jørgensen, A. Geertz, and A. Roepstorff (2008) Rewarding prayers. Neuroscience Letters 443 (2008) 165–168.
D. Westen, P. Blagov, K. Harenski, C. Kilts, S. Hamann. (2006) Neural Bases of Motivated Reasoning: An fMRI Study of Emotional Constraints on Partisan Political Judgment in the 2004 U.S. Presidential Election. Journal of Cognitive Neuroscience. 18: 1947-1958.
Charles Brack