God, Meet Darwin
Religiosity, Reproductive Advantage, and Sexual Behavior
How did these two adversaries end up on the same side?
Of the 6.5 billion humans on this planet, roughly 8% have no spiritual or religious beliefs, which poses the interesting question: what evolutionary business does this subpopulation of non-believers have with the believers? To date, the limited genetic evidence implies that the probability of inheriting your religious disposition from your parents is just about the same as inheriting your intelligence. While environmental influences weigh heavily on religious disposition, the evidence indicates that genetics, on average, is more important (see Brouchard et al. 1990).
But what genes? The number of genes contributing to religious (and political) disposition might number in the thousands, but given some recent evidence, those genes associated with the development and functioning of the monoamine neurotransmitter systems, in particular the dopaminergic system, seem to exert a disproportionate influence on the modulation of religiosity and political conservatism.
But when you talk about genetic inheritance and inter and intraspecies competition, you are talking about natural selection. This raises the question: how much has religion impacted the frequencies of certain genes, such as the dopamine D4 receptor gene (DRD4)? The impact of the so-called "adventure" DRD4 gene on human social behavior has attracted a lot of interest, as it has been connected to excessive gambling, substance abuse, elevated levels of novelty seeking, and the acceptance of spiritual transcendence. However, DRD4 studies have led to contrasting results, as the isolation of single genes to explain aggregate behaviors is a risky business.
But, in one of the greatest ironies in human evolutionary history, religiosity has become an active agent in the evolution of the human nervous system. In a sense, religiosity and the genes that promote it have formed a runaway selective process analogous to Fisher's runaway sexual selection. As we shall see, the religious are the most piously discerning when it comes to selecting a mate, that is, they heavily apply religious criteria towards potential mates in what we believe to be a Darwinian attempt to improve their reproductive output.
We suspect that the confluence of those behaviors promoted by religious doctrine combined with the genes that promote religiosity are a runaway evolutionary process. The behaviors promoted by religions tend to facilitate reproductive output, and further, provide a cultural blanket that protects the genes that promote religiosity. The most conspicuous cultural aspect of religions is morality, which we have argued to promote both reproductive output and advantage, along with behavioral inhibition of socially inappropriate reward-seeking.
The cultural-religious importance of morality lies in the fact that it is primarily targeted towards regulation of the behaviors associated with the dopamine system, known for its predisposition towards reward-seeking. In short, morality is a system of checks and balances for the human dopamine system. The religious do not focus on morality because they want to. They focus on it because the have to, that is, it facilitates higher reproductive output.
Empathy-based controls on reward-seeking are the domain of the secular liberal, and the neural substrates supporting empathy are not as dependent on the dopamine system as is morality, and are more likely to incorporate the noradrenergic system. The moral framework associated with religion provides the social controls to offset the dopaminergic nature of religious disposition. The religious do not, and can not, rely on empathy in regulating their social behavior to the extent that the secular liberals can. Indeed, religions transfer earthly reward-seeking into the afterlife, where heavenly rewards await those that inhibit their anti-social and selfish behavior on earth.
But the religious moral framework, combined with the dopaminergic nature of religious disposition, has led to several substantial Darwinian advantages. These are:
1.
Reproductive yield
2. The survival of small breeding populations
3. Emigration to relieve population stress
4. Economic collusion
5. Welfare and income redistribution
All of the above advantages are interrelated. We shall briefly touch on the survival of small breeding populations and emigration, but the focus of this month's edition will be reproductive output and reproductive advantage. The evidence for elevated reproductive output by degree of religiosity is substantial. This is illustrated in the graph below.
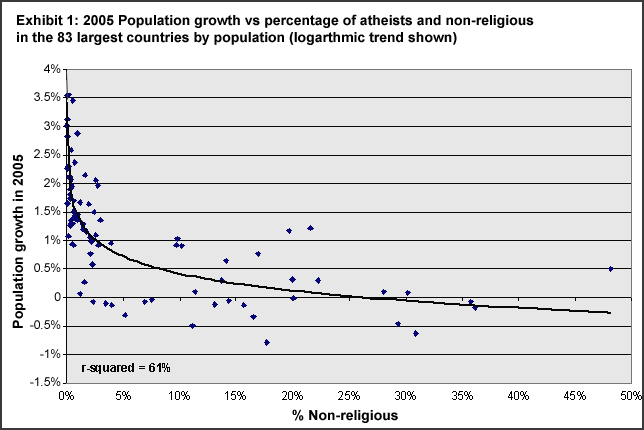
2005 Population growth by percentage of non-religious
Source: World Christian Database
The percentage of religious in a population is highly correlated with reproductive rates. Further, reproduction is highly correlated with the degree of religiosity. So how are the religious doing it? What is the secret to their Darwinian success?
A, B, and C
Among humans, reproductive strategies are heavily integrated with social alliances, which collectively form the foundation for religious and political behavior. Due in part to the higher reproductive rates of the religious and the mounting evidence of the genetic heritability of religious disposition, natural selection has guided the human gene pool to its current religious bias.
In spite of the reproductive advantages of religiosity, agnosticism and atheism are still prominent tendencies, which should probably be interpreted as an evolutionary complement to religiosity. That is, a certain level of non-religiousness in a population actually improves the reproductive yield of the religious, as those with occupations in science and technology tend towards lower rates of religiosity. Humans may indeed be in an evolutionary stalemate between the genes that facilitate religiosity and reproduction, and the genes that improve technological advancement and the carrying capacity of habitats.
While a certain percentage of any large population is nonreligious, there is no current evidence that this percentage remains stable over time. Instability in this percentage is further promoted by environmental influences, which can rapidly modulate levels of religiosity. Further, population density and religiosity are inversely correlated variables.
Religious groups are a prominent fixture in the history of the colonization of new territories, and their high levels of group altruism and tight social organization provide an effective model for the survival and fertility of small populations in unfriendly habitats. In population biology, the religious most closely resemble the behavioral and reproductive characteristics of r-selected animal populations.
The elevation in reproductive output by the religious is executed by a diverse combination of sexual behaviors, fertility stimulants, and parenting strategies. In this light, we shall present our data in the following categories:
1. Desire for offspring, religious and racial tendencies in mate selection
2. Active relationships, relationship duration, numbers of different sex
partners, and monogamy
3. Non-reproductive sexual behaviors
4. Male dominance, orgasmic ability, and masturbation
Methods and Data
The data represented here was collected from three separate surveys consisting of 7,488 completed question-naires administered from March 2006 through November 2007. Religious disposition in these surveys was self-rated. For correlation analysis, these religiosity cohorts were assigned a numerical religiosity variable, on a scale of 0 to 5, with atheistic = 0, agnostic = 1, spiritual = 2, little religious = 3, moderately religious = 4, and the very religious = 5.
All correlation coefficients are Pearson coefficients. For certain responses, which specified a range instead of a single value, we used the midpoint of that range in statistical analysis. If the top range was unbounded, (e.g. 100+ sex partners), 100 was used instead as the midpoint. This under estimates the calculation of the mean, and we have noted these statistics with an asterisk "*".
The data is summarized into two different schemas: religious, spiritual, and nonreligious, and, by degree of religiosity or nonreligiosity. These degrees will be atheist (AT), agnostic (AG), very religious (VR), moderately religious (MR), and a little religious (LR).
The religious cohort will include the very religious, moderately religious, and little religious. The nonreligious cohort will include the agnostics and atheists. The spiritual will not be aggregated into either the religious or nonreligious cohorts, although their responses are more likely to resemble the nonreligious.
1. Desire for Offspring, Religious and Racial Tendencies in Mate Selection
Mate selection is a critical element in reproductive yield, as mismatches in the number of desired children can reduce the reproductive output of the partner with the greatest desire. Since desire for offspring varies with the level of religiosity, selecting partners for religiosity has the interesting side-effect of also matching the desire for children. Indeed, this seems to be the Darwinian secret underlying the strong tendency for the religious to seek out religious partners.
|
Gender |
Religious Disposition |
N |
Mean Number of Children Desired* |
N |
Religious Affiliation Match with
Partner |
N |
Racial Match with Partner |
|
Female |
Religious |
250 |
2.6 |
404 |
54.7% |
340 |
83.2% |
|
|
Nonreligious |
252 |
1.7 |
322 |
34.5% |
264 |
85.6% |
|
|
Spiritual |
181 |
2.0 |
212 |
42.0% |
180 |
80.6% |
|
|
VR |
48 |
3.0 |
135 |
68.2% |
108 |
85.2% |
|
|
MR |
115 |
2.4 |
172 |
51.7% |
146 |
82.9% |
|
|
LR |
87 |
2.5 |
97 |
41.2% |
86 |
81.4% |
|
|
AG |
126 |
1.8 |
238 |
31.9% |
199 |
85.9% |
|
|
AT |
126 |
1.6 |
84 |
41.7% |
65 |
84.6% |
|
Male |
Religious |
499 |
2.7 |
982 |
57.5% |
802 |
84.2% |
|
|
Nonreligious |
630 |
1.7 |
791 |
24.3% |
564 |
80.3% |
|
|
Spiritual |
206 |
2.2 |
311 |
32.8% |
234 |
78.2% |
|
|
VR |
134 |
3.5 |
306 |
64.7% |
250 |
86.8% |
|
|
MR |
226 |
2.5 |
459 |
58.8% |
379 |
85.8% |
|
|
LR |
139 |
2.1 |
217 |
44.7% |
173 |
76.9% |
|
|
AG |
226 |
1.9 |
500 |
24.8% |
356 |
80.6% |
|
|
AT |
404 |
1.6 |
291 |
23.4% |
208 |
79.8% |
(AT=Atheist, AG=Agnostic, LR=A little religious, MR=Moderately religious, VR=Very religious) (Religious=VR+MR+LR, Nonreligious=AG+AT)
The number of children desired is correlated with religiosity, as evident in Table 1. The very religious (VR) in both genders had the highest mean preference for offspring, with females desiring 3.0 children, and males 3.5. These averages are about twice as high as atheists (AT), with females and males desiring 1.6 children.
The religious, in the process of seeking mates with common religious affiliations, are effectively seeking mates that match their desire for children and their reproductive strategies in general. This is evident from the higher correlation of partner religious affiliation with the level of religiosity (r=0.29, p<0.0001). The religious affiliation options in the questionnaire included Christian, Catholic, Mormon, Judaism, Buddhism, Hinduism, Islam, Confucianism, spiritual, agnostic, atheist, and other. If the survey respondent had a partner, we only considered it a match if the partner's religious affiliation matched the respondent's (based on the aforementioned categories). For example, it would only be considered a match if an atheist had an atheist partner, and if someone was Catholic, it would not be a match if their partner was Christian.
The very religious (VR) had the highest rate of matching their religious affiliation with their partners (females = 68.2%, males = 64.7%). Among males, the atheists (AT) had the lowest rate of matching religious affiliation (23.4%), while among the females; the agnostics (AG) had the lowest rate of matching (31.9%).
Seeking partners with the same religious affiliation is most important for those with a greater desire for offspring (i.e., the very religious), as selecting a mate with a desire for fewer children would tend to reduce the probability of achieving one's reproductive goals. Indeed, the religious were more likely to have an opinion about their partner's religion, and the nonreligious were more likely to be unsure.
This same phenomenon is also seen in partner racial assessments. The religious are more likely to have an opinion about their partner's race, and among the males, the religious are more likely to indicate they are of the same race as their partner. We asked the survey respondents to classify themselves into one of the following racial groups: Asian, Black, Hispanic, Indian, Middle Eastern, Native American, and White. "Not sure" was also an option in defining partner race.
For females, there was no correlation between religiosity and racial matching. However, for males, it was correlated (r=0.13, p<0.0001), as the very religious (VR) and moderately religious (MR) males are substantially elevated when it comes to matching the race of their partner (86.8% and 85.8% respectively). Conversely, the male atheists (AT) and agnostics (AG) were the least likely to match their partner's racial group (79.8% and 80.6%).
The females in this survey were more likely to have an opinion about their partner's race, and to also report they were of the same race. While the female tendency towards racial sensitivity in mate selection can be attributed to the greater female investment in reproduction, what evolutionary game are the religious males playing? Perhaps it is the same game, that is, the elevated investment in reproduction corresponds with greater sensitivity to the reproductive attributes of females.
2. Active Relationships, Relationship Duration, Numbers of Different Sex Partners, and Monogamy
Species with lower reproductive rates tend to neutralize their reproductive deficit by enhanced parental investment strategies. As a slowly reproducing species, humans have evolved an interesting spectrum of such strategies, and religiosity is an active component. In table 2, we see four variables with overtones in parental investment and reproductive strategies in general.
Religiosity and the probability of being in an active relationship are correlated variables in both genders. An active relationship in this context simply means that the person indicated they currently have a partner, although we did not identify whether this relationship was heterosexual or not. Among females, having an active partner varied with the level of religiosity.
|
Gender |
Religious Disposition |
N |
Percent in an active relationship |
Mean length of relationship (in
years)* |
Mean different sex partners* |
Self-rated Monogamy |
|
Female |
Religious |
250 |
84.8% |
8.4 |
10.1 |
9.2 |
|
|
Nonreligious |
252 |
77.4% |
6.6 |
14.1 |
8.5 |
|
|
Spiritual |
181 |
75.1% |
7.5 |
16.2 |
8.7 |
|
|
VR |
48 |
89.6% |
10.3 |
5.6 |
9.6 |
|
|
MR |
115 |
84.4% |
8.1 |
10.3 |
9.4 |
|
|
LR |
87 |
82.8% |
7.6 |
12.4 |
8.6 |
|
|
AG |
126 |
79.4% |
7.2 |
13.3 |
8.7 |
|
|
AT |
126 |
75.4% |
6.1 |
14.9 |
8.3 |
|
Male |
Religious |
499 |
79.0% |
11.1 |
13.3 |
8.8 |
|
|
Nonreligious |
630 |
62.9% |
7.2 |
14.7 |
8.2 |
|
|
Spiritual |
206 |
67.5% |
8.4 |
19.4 |
8.3 |
|
|
VR |
134 |
75.4% |
12.7 |
9.3 |
9.3 |
|
|
MR |
226 |
78.8% |
11.1 |
14.3 |
8.8 |
|
|
LR |
139 |
82.7% |
9.5 |
15.3 |
8.4 |
|
|
AG |
226 |
65.9% |
7.4 |
15.8 |
8.3 |
|
|
AT |
404 |
61.1% |
7.1 |
14.1 |
8.2 |
(AT=Atheist, AG=Agnostic, LR=A little religious, MR=Moderately religious, VR=Very religious) (Religious=VR+MR+LR, Nonreligious=AG+AT)
Male religiosity was more strongly correlated with the probability of being in an active relationship (r=0.15, p<0.0001) than was female religiosity (r=0.09, p<0.02). The little religious males reported the highest rate, at 82.7%, while the very religious females recorded the highest overall rate (89.6%). The atheist males and females reported the lowest rates of active relationships, at 61.1% and 75.4% respectively. In this survey, females were more likely to report an active relationship, although we did not inquire as to how many active relationships the individual was currently a part of.
Looking solely at the people reporting an active relationship, we computed their mean relationship length. The trends are again pronounced, with the length of the active relationship varying with the level of religiosity, in both genders. The very religious males had the longest relationship length, at 12.7 years, followed immediately by the moderately religious males at 11.1. The atheistic males had the shortest average duration, at 7.1 years.
While females reported shorter relationship durations on average, they mirrored the male pattern quite well, in which length varied with strength of religiosity. Long relationships support the reproductive investment necessary to accommodate the greater desire for offspring, facilitating a high-yield reproductive strategy.
We also computed the average number of different sex partners for our religious-gender cohorts. Among females, religiosity was negatively correlated with the number of sex partners (r=-0.12, p<0.002). The very religious females reported the lowest number (5.6), and the moderately religious were next (10.3). The atheistic (14.9) and agnostic (13.3) females reported higher numbers, but not nearly as high as the spiritual (16.2).
Among males, the negative correlation between religiosity and numbers of sex partners was weaker, (r=-0.04, p<0.16). The outliers for the number of sex partners were the very religious (9.3) and the spiritual (19.4). Like the spiritual females, the spiritual males led their gender in numbers of different sex partners, an interesting result indeed.
When it comes to reproduction, the spiritual resemble the atheists and agnostics more than they do the religious: one of the reasons we have excluded them from the religious cohort. People that define themselves as spiritual, yet have no religious affiliation, differ substantially from the religious on a large number of cognitive and behavioral attributes.
Finally, monogamy was self-rated, on a scale of 1 to 10, with 10 being the highest level. The highest ratings were among the very religious in both genders (males = 9.3, females = 9.6). The moderately religious in both genders were next, with males averaging 8.8 and females averaging 9.4. The agnostics and atheists in both genders had the lowest self-ratings of monogamy. The level of religiosity and self-rated monogamy were closely correlated variables in both genders: females (r=0.18, p<0.0001), males (r=0.16, p<0.0001).
3. Non-reproductive Sexual Behaviors
One of the prominent variations between the religious and nonreligious is the stronger reproductive valence maintained by the religious in their sexual behavior. A sample of sexual behaviors that do not directly lead to conception are displayed in table 3, which includes self-reported arousal levels for anal sex, oral sex, pornography, and homosexual experience.
Arousal levels for anal and oral sex were based on a scale of 1 to 10, with 10 being the highest level of arousal one could report. However, the questions were not framed to delineate whether one was aroused by giving or receiving oral or anal sex. For viewing pornography, the respondent was asked to indicate one of four options available: never, rarely, sometimes, and frequently. In table 3, only those responding sometimes and frequently were included in the statistics. For homosexual experience, we combined the rarely, sometimes, and frequently responses in this statistic, and excluded those indicating they never had a homosexual experience. Note that in this survey, the experience should have resulted in an orgasm to be classified as homosexual.
|
Gender |
Religious Disposition |
N |
Mean arousal for anal sex |
Mean arousal for oral sex |
Percentage looking at pornography
(frequently or sometimes) |
Percent having at least one
homosexual experience |
|
Female |
Religious |
250 |
4.2 |
5.9 |
36.4% |
18.8% |
|
|
Nonreligious |
252 |
4.9 |
6.1 |
46.0% |
32.5% |
|
|
Spiritual |
181 |
4.7 |
6.3 |
46.4% |
34.3% |
|
|
VR |
48 |
2.8 |
5.0 |
14.6% |
10.4% |
|
|
MR |
115 |
4.3 |
6.2 |
39.1% |
18.3% |
|
|
LR |
87 |
4.8 |
5.9 |
44.8% |
24.1% |
|
|
AG |
126 |
4.6 |
6.1 |
49.2% |
29.4% |
|
|
AT |
126 |
5.2 |
6.1 |
42.9% |
35.7% |
|
Male |
Religious |
499 |
5.4 |
7.2 |
70.7% |
16.4% |
|
|
Nonreligious |
630 |
6.3 |
7.6 |
84.3% |
23.2% |
|
|
Spiritual |
206 |
6.3 |
7.8 |
77.7% |
23.3% |
|
|
VR |
134 |
5.0 |
7.1 |
49.3% |
15.7% |
|
|
MR |
226 |
5.3 |
7.2 |
76.6% |
15.9% |
|
|
LR |
139 |
5.9 |
7.4 |
82.0% |
18.0% |
|
|
AG |
226 |
6.3 |
7.6 |
85.0% |
22.1% |
|
|
AT |
404 |
6.3 |
7.6 |
83.9% |
23.8% |
(AT=Atheist, AG=Agnostic, LR=A little religious, MR=Moderately religious, VR=Very religious) (Religious=VR+MR+LR, Nonreligious=AG+AT)
The males were more aroused by anal sex than females, but the very religious were again at the bottom of the male scale (5.0). Similar to the female pattern, the moderately religious males were next at 5.3. The atheists, agnostics, and spiritual males all reported the same level of arousal, at 6.3, yet another statistic where the spiritual resemble the nonreligious more than the religious.
Arousal for oral sex was substantially higher in both genders, and indeed there is some evidence that oral sex may help prevent preeclampsia (see Koelman et al. 2000). But religiosity was inversely correlated with arousal for oral sex in both genders, and the very religious reported the lowest arousal scores (females = 5.0, males = 7.1). Interestingly, the spiritual reported the highest rates of arousal in both genders (females = 6.3, males = 7.8), another indicator of the variation between the religious and the spiritual. Among females, outside of the very religious, the correlation was weak between religiosity and arousal to oral sex (r=-0.05, p<0.23).
Finally, religiosity was an active trait in the reporting of homosexual experiences (to the point of orgasm). The greater the level of religiosity, the less likely one will report having a homosexual experience. The very religious reported the lowest rates (females = 10.4%, males = 15.7%), while the spiritual (females = 34.3%, males = 23.3%) and the atheists (females = 35.7%, males = 23.8%) reported the highest rates.
Males, in general, were more likely to indicate they were dominant in their relationships. In this relationship dominance statistic, we only included those in an active relationship. Among females, relationship dominance is negatively correlated with the level of religiosity (r=-0.05, p<0.21), while among males, it is positively correlated (r=0.09, p<0.01). We must note that the heterosexual or homosexual nature of these relationships was not differentiated in this statistic.
While the religious indicated an elevation in copulatory orgasmic ability, the nonreligious males were still effective in the ability to achieve orgasms during copulation. 90.9% of the nonreligious males always or frequently achieved orgasm during intercourse, compared to 91.2% of the religious males and 89.7% of the spiritual males.
But
among females, the nonreligious reported a deficit in orgasmic ability, as
43.9% reported they always
or frequently
achieved orgasm during intercourse, compared to 50.8% of the religious and
48.5% of the spiritual. Note that the
very religious females were the most orgasmic during intercourse, at 58.7%.
The interesting elevation of female orgasmic ability among the religious
(during intercourse) might be evidence for a reproductive function to the
female orgasm; however, we will defer this complex and controversial topic
until later, as this can actually be interpreted to support several different
theories of the evolutionary function of the female orgasm, such as Symons
(1979).
|
Religious
Disposition |
N |
Percentage indicating they are dominant in relationship |
Orgasms
easier with intercourse than with masturbation |
Orgasm
during intercourse (frequently or always) |
Mean
Masturbation per week* |
|
Religious |
250 |
23.6% |
24.4% |
50.8% |
1.8 |
|
Nonreligious |
252 |
26.2% |
19.8% |
43.9% |
2.2 |
|
Spiritual |
181 |
24.6% |
20.8% |
48.5% |
2.3 |
|
VR |
48 |
13.0% |
25.0% |
58.7% |
1.6 |
|
MR |
115 |
27.0% |
21.6% |
48.3% |
1.7 |
|
LR |
87 |
24.7% |
27.7% |
50.0% |
2.1 |
|
AG |
126 |
22.9% |
20.7% |
38.8% |
2.2 |
|
AT |
126 |
29.4% |
19.0% |
49.1% |
2.2 |
|
Religious |
499 |
39.9% |
51.3% |
91.2% |
3.2 |
|
Nonreligious |
630 |
34.3% |
37.2% |
90.9% |
4.4 |
|
Spiritual |
206 |
36.5% |
41.6% |
89.7% |
4.2 |
|
VR |
134 |
44.4% |
61.9% |
94.9% |
2.9 |
|
MR |
226 |
41.0% |
49.1% |
90.2% |
3.2 |
|
LR |
139 |
33.9% |
45.4% |
89.5% |
3.7 |
|
AG |
226 |
34.3% |
39.6% |
88.1% |
4.3 |
|
AT |
404 |
34.3% |
36.0% |
92.4% |
4.5 |
(AT=Atheist, AG=Agnostic, LR=A little religious, MR=Moderately religious, VR=Very religious) (Religious=VR+MR+LR, Nonreligious=AG+AT)
Finally, masturbation is negatively correlated with religiosity in both genders (females: r=-0.09, p<0.02; males: r=-0.22, p<0.0001). The reproductive implications for male masturbation relate primarily to semen volume, sperm count, motility, and morphology (Pellestor et al., 1994), and imply an interesting trade-off between sperm volume and sperm quality, dependent on ejaculation frequency. The lower rates of religious male masturbation might imply that they are releasing more sperm during copulation; an advantage in fertility, especially when combined with the lowered tendency for the religious to use contraception (Studer and Thornton, 1987).
Discussion
By some accounts, Joseph Smith, the founder of Mormonism, had as many as 84 plural marriage partners (Compton, 1997), which highlights the interesting relationship between cult formation and reproductive advantage. The reproductive advantages of cult founders can be substantial (Galanter, 1989), as seen more recently with David Koresh and his preferential sexual access to females within the Waco Branch Davidian compound (C. Wilson, 2000).
Primate studies indicate that dominant males maintain substantial reproductive advantages over subordinate males, especially when the number of males in the group is small (Colishaw and Dunbar, 1991). As the number of males increase, the advantages of dominance are neutralized by the costs incurred to guard mates and monopolize sexual access. However, organized cults frequently employ behavioral regulation mechanisms that maintain the reproductive advantages of cult leaders, along with emigration and colony formation to reduce the potentially disruptive influences of kin.
The relationship between religiosity and emigration is interesting, as there are many parallels with the definition of r-strategists in the field of population biology. r-strategists migrate to new habitats quickly, reproduce rapidly, and disperse rapidly when those habitats are depleted. They are noted by elevations in several altruistic traits, such as mutual defense, cooperative foraging, and nest building (E. Wilson, 2000). Extinctions of small colonies are highest at two points: at the beginning of colonization; and at the point the population has reached the maximum carrying capacity of its habitat.
Religiosity has a positive impact on colonial longevity, as documented in the Sosis and Bressler study (2003) of 19th century communes. The religious colonies in this study had a mean longevity of 35.6 years, while the secular colonies lasted 7.7 years. Sosis noted that intragroup conflict and economic failure were the two most common reasons for commune dissolution. Sosis’ costly signaling theory of religion proposes that the greater the behavioral regulations placed on the members, (e.g., celibacy and fasting), the greater the longevity of the commune. However, as Sosis noted, there was no relationship between “costly signaling” and longevity in secular communes, creating a significant issue with the theory. Sosis also noted that religious communes were more likely to organize around a charismatic leader than were secular communes.
The relationship between religiosity and successful colonization is no doubt the result of the highly coordinated social behavior and mutual altruism practiced by organized religious groups, but of the eighty-three 19th century communes studied by Sosis, only a single religious commune has remained active until today--the Hutterites.
The ability of organized religious groups to withstand r-extinctions, or extinctions of new colonies, is certainly implied by the Sosis study. A primary difference between colonies formed by cults and colonies formed by existing religious sects lies in the level of support from the parent population, either through kinship or religious ties. Cults are more likely to isolate themselves, thereby sustaining the dominance and reproductive advantages of the cult leaders.
But from a perspective of population biology, there are several factors that have a significant impact on the total reproductive output of a colony (or any population, for that matter), such as the numbers of females and males that are in an active relationship that supports reproduction. Based on the results in table 2, the religious males have a substantial advantage over the nonreligious males, and this implies a substantial reproductive advantage.
This poses an interesting question: does religiosity improve the likelihood that a male will reproduce? Across most sexually-reproducing species, the percentage of females that produce at least one offspring is greater than the percentage of males. Religiosity in males seems to improve the percentage that sire offspring.
The religious also report higher levels of monogamy and longer relationship durations. This should ordinarily work to improve reproductive yield, especially in cases where both parties desire children. Interestingly, the desire for the religious to have religious partners, as noted in table 1, seems to be facilitating the matching of the desire for offspring, and ultimately reproductive output.
The correlation between religiosity and male dominance, as noted in table 4, pertains to dominance within the male-female relationship, and is not directly associated with intrasexual, or male-male dominance. The correlation between intrasexual male dominance, sexual dimorphism, and reproductive advantage has certainly been documented in numerous species, but how does male dominance over females impact reproductive rates?
While the Gottschall study (2003) indicates that fertility rates were higher in cases of rape than with consensual sex, fertility rates pertaining to varying degrees of sexual coercion among monogamous couples is unknown. But we can speculate that male dominance within a relationship improves sexual access by the male. Following this line of reasoning, the very religious males, reporting the highest level of dominance within their relationships, would have greatest sexual access to their partners. The fertility implications are significant, as they also desire more children than any other cohort.
This brings us to the issue of the reported elevation in orgasmic ability among the religious females. The connection between male dominance and female orgasmic ability would certainly be an expected outcome of natural selection, and indeed, it has been documented with other species (Troisi and Carosi, 1998). How this relates to the more submissive role played by religious females is speculative, but the Donald Symons’ hypothesis of the female orgasm as an inert embryological byproduct, like the male nipple, would be challenged by the fact that females that are more reproductive are also more likely to orgasm during intercourse. This is not to imply that the act of female orgasm improves fertilization specifically, rather, it seems to facilitate the general process of reproduction. The female orgasm would presumably provide a substantial reward to induce copulation, along with the neurological chain of events that promote romantic bonding, ultimately improving reproductive yield.
The proposal that female orgasmic ability is an evolutionary adaptation that promotes female choice in mate selection may have some merit when applied to the religious. However, mate selection may only be part of the orgasmic advantage, as long-term bonds seem to be facilitated by long-term copulatory orgasmic ability. Note that the religious females reported longer average relationships, and remarkably, higher rates of copulatory orgasmic ability in those relationships.
But perhaps the most significant variable associated with the reproductive advantage is the desire for children. As seen in table 1, this is strongly correlated with religiosity. While the religious seem to be playing a very tactical game of reproductive advantage, they are, for the most part, unaware of this game. Evolution has loaded the nervous system with simple, yet effective, sexual and nonsexual adaptations that promote reproduction.
One of these adaptations is evident in the table below, where the survey respondents were asked to select a favorite odor from the following options: chocolate, coffee, vanilla, garlic, or a newborn baby. There is indeed a positive correlation between religiosity and the hedonic preference for the odor of a newborn baby in both genders, (r=0.11, p<0.0001), which could be interpreted as an evolutionary adaptation that improves reproductive yield.
|
Gender |
Religious
Disposition |
N |
Newborn
Baby |
Coffee |
Chocolate |
Garlic |
Vanilla |
|
Female |
Religious |
335 |
17.0% |
31.0% |
16.7% |
7.2% |
28.1% |
|
|
Nonreligious |
381 |
8.7% |
38.3% |
16.5% |
12.1% |
24.4% |
|
|
Spiritual |
229 |
14.9% |
29.7% |
14.9% |
13.1% |
27.5% |
|
|
VR |
81 |
24.7% |
24.7% |
21.0% |
6.2% |
23.5% |
|
|
MR |
156 |
16.0% |
30.8% |
16.7% |
7.1% |
29.5% |
|
|
LR |
98 |
12.2% |
36.7% |
13.3% |
8.2% |
29.6% |
|
|
AG |
258 |
9.7% |
38.8% |
14.3% |
11.6% |
25.6% |
|
|
AT |
123 |
6.5% |
37.4% |
21.1% |
13.0% |
22.0% |
|
Male |
Religious |
567 |
7.8% |
36.9% |
15.9% |
10.8% |
28.8% |
|
|
Nonreligious |
672 |
3.3% |
39.7% |
13.1% |
11.8% |
32.1% |
|
|
Spiritual |
264 |
7.6% |
43.6% |
12.9% |
9.9% |
26.1% |
|
|
VR |
145 |
13.1% |
33.8% |
12.4% |
12.4% |
28.3% |
|
|
MR |
245 |
6.1% |
37.6% |
18.0% |
10.6% |
27.8% |
|
|
LR |
177 |
5.7% |
38.4% |
15.8% |
9.6% |
30.5% |
|
|
AG |
257 |
3.1% |
42.4% |
12.8% |
10.6% |
31.1% |
|
|
AT |
415 |
3.5% |
35.4% |
13.6% |
13.6% |
33.9% |
(AT=Atheist, AG=Agnostic, LR=A little religious, MR=Moderately religious, VR=Very religious) (Religious=VR+MR+LR, Nonreligious=AG+AT)
However, in this response, we cannot be sure that they were solely influenced by the odor cues associated with infants, and not some other sensory feature. The neurobiology of parent-child bonding is complex, and involves the phylogenetically ancient olfactory system. If indeed the hedonic preference for the odor of a newborn baby is genetically modulated, it is only one of many evolutionary adaptations that improve reproduction.
It is also interesting to note that this preference is one of the few instances where the spiritual resemble the religious. The spiritual, or those that have a belief in a deity, yet no religious affiliation, actually look more like the atheists and agnostics in their questionnaire responses. Perhaps this unusual cognitive alliance between the spiritual and the nonreligious is explained by hemisphericity, or the various mechanisms employed by the brain to resolve conflicts in the alternate behaviors and cognition promoted by the lateralization of the left and right cerebral hemispheres.
The
brain is a paired-organ; with each half specialized in cognitive and behavioral
functionality (Demaree et al., 2005). The left cerebral hemisphere is implicated with
the propensity to apply single meanings to ambiguous targets, while the right
hemisphere is prone to apply alternate meanings to the same targets (Faust
and Lavidor, 2003). The general mode of left hemispheric cognition seems to
be the reduction of ambiguity, which has been associated with political conservatism
(Jost et al., 2003), which itself is correlated with religiosity.
There
is an interesting convergence along several different lines of neurocognitive
research with implications for a special role of the left cerebral hemisphere
in religiosity and political conservatism.
The first comes from the study of belief-bias, which has identified
a left temporal lobe system in belief-biased reasoning, while the right lateral
prefrontal cortex is engaged when subjects inhibit a prepotent response associated
with belief-bias (Goel and Dolan, 2003).
The
assignment of people, objects, and events into “good” or “evil” categories
is a recurrent theme in religious dogma, and the neurology supporting this
assignment appears to be centered in the left parietal lobe (Newberg et al.,
2001). This region is also implicated in motor planning
for the redirection of body movements, which may hint that the binary categorization
of “good” and “evil” has its evolutionary origins in the neurology associated
with the fight-or-flight instinct.
Within
the left temporal lobe, asymmetrically large hippocampal volumes are correlated
with higher levels of religiosity and refractory epilepsy (Wuerfel, 2004),
further implicating the left cerebral hemisphere’s role in religious disposition.
But perhaps the most significant neurological relationship to religiosity
lies with the dopaminergic system (Previc, 2006). Also, see last month's
edition. The dopaminergic system is an anatomically distinct arrangement
of neural pathways that activate cortical and sub-cortical structures to facilitate
motor-control, reward-anticipation, reward-seeking, and approach.
The
dopaminergic system, implicated in romantic bonding, sexual function, meditative
states (Kjaer et al., 2002), and reward prediction, makes it a prime candidate
in the behaviors associated with religiosity and political conservatism.
The reproductive rates of the religious are better supported by a higher
propensity for reward-seeking; however, to date, there is no direct evidence
that the religious are elevated in their propensity for reward seeking.
With
the reproductive advantages of religiosity, why does agnosticism and atheism
persist? This answer to this may lie
in the social integration of divergent cognitive styles, that is, optimization
of one’s contributions and compensations within a social group occurs when
individual brains are specialized for different sets of tasks. This phenomenon is seen in vertebrates (Vallortigara,
2000), and our data pertaining to occupation indicates that the religious
and nonreligious are performing different economic tasks. The religious and
nonreligious seem to have developed a rather symbiotic division of labor,
forming a sort of economic glue that counteracts their tendency for social
and political conflict.
In
the aforementioned Sosis study of 83 nineteenth century religious and secular
communes, 82 failed, following the general tendency of high extinction rates
for small animal populations. Over
the same time period, the survival rate of cities with populations of over
one million was one-hundred percent.
Extreme levels of religiosity would seem to have an evolutionary cost, as the time and effort devoted to religious behavior leaves less time for technological and economic pursuits. Indeed, secularism and technological change are highly correlated variables, and reproductively successful religious populations must maintain some level of integration with the economics, science, and technology of dynamic secular populations, while minimizing their negative impact on fertility.
It is certainly arguable that secularism, via a greater tendency for cognitive diversity, specialization, and technological change, has actually improved the carrying capacity of habitats, subsequently supporting larger religious populations. Ironically, atheism and agnosticism may indeed be an evolutionary complement to religiosity, and the evolutionary trajectory of human gene pool seems to be walking the fine line between reproductive effectiveness and technological progress.
The
strong orientation of the religions toward the afterlife, morality, male dominance,
heterosexuality, fidelity, etc., when combined with the genes that promote
religiosity, is fundamentally a runaway
References:
Bouchard et al. Sources
of human psychological differences: the Minnesota Study of Twins Reared
Apart. Science 12 October 1990: 223-228
G. Colishaw and R. Dunbar.
Dominance rank and mating success in male primates. Animal Behavior.1991.
41, 1045-56
T. Compton, In Sacred
Loneliness: The Plural Wives of Joseph Smith. 1997. Introduction.
Signature Books
H. Demaree, E. Everhart,
E. Youngstrom, and D. Harrison. Brain Lateralization of Emotional
Processing: Historical Roots and a Future Incorporating "Dominance".
Behavioral and Cognitive Neuroscience Review. 2005;4;3
M. Faust and M. Lavidor. Semantically convergent and semantically
divergent priming in the cerebral hemispheres: lexical decision and
semantic judgment. Brain Research. Cognitive Brain Research. 2003
Oct;17(3):585-97.
M. Galanter. Cults and
New Religious Movements. 1989. American Psychiatric Association.
V. Goel and R. Dolan. Explaining modulation of reasoning by belief.
Cognition 87. 2003.
J. Gottschall and T. Gottschall.
Are per-incident rape-pregnancy rates higher than per-incident consensual
pregnancy rates? Human Nature. March 2003. Vol 14, No 1.
J. Jost, J. Glaser, A. Kruglanski, and F. Sulloway. Political Conservatism
as Motivated Social Cognition. Psychological Bulletin. 2003. Vol.
129, No. 3. p. 339-75.
T. Kjaer et al. Increased dopamine tone during meditation-induced
change of consciousness. Brain Research. Cognitive Brain Research.
Apr 2002; 13(2):255-9.
CA Koelman et al. Correlation between oral sex and a low incidence
of preeclampsia: a role for soluble HLA in seminal fluid? Journal
of Reproductive Immunology, March 2000; 46(2): 155-66.
A. Newberg, E. D'Aquili,
and V. Rause. Why God Won't Go Away. 2001. Ballantine Publishing Group.
F. Previc. The Role of Extrapersonal Brain Systems in Religious
Activity. Consciousness and Cognition. 2006.
R. Sosis and E. Bressler.
Cooperation and Commune Longevity: A Test of the Costly Signaling
Theory of Religion. Cross-Cultural Research. May 2003. Vol 37, No.
2, p.211-39
M. Studer and A. Thornton,
Adolescent Religiosity and Contraceptive Use. 1987. Journal of Marriage
and the Family. 49:117:28.
D. Symons. The Evolution of Human Sexuality. Oxford University Press.
1979.
Pellestor et al. Effect of long abstinence periods on human sperm
quality. Int J of Fertility and Menopausal Studies. 1994 Sep-Oct.;
39(5):278-82.
A. Troisi and M. Carosi. Female orgasm rate increases with male dominance
in Japanese macaques. Animal behavior. November 1998. Vol 55. Issue
5.
D. Tucker and P. Williamson. Asymmetric Neural Control Systems in
Human Self-Regulation. Psychological Review, 1984, 91, 185-215.
G. Vallortigara. Comparative
Neuropsychology of the Dual Brain: A Stroll through Animals' Left
and Right Perceptual Worlds. Brain and Language. June 2000. p 189-219
C. Wilson. The Devil's
Party. 2000 Virgin Books.
E. Wilson. Sociobiology. 2000. Belknap Press. p.99-103, 107
J. Wuerfel et al. Religiosity is associated with hippocampal but not
amygdale volumes in patients with refractor epilepsy. Journal of Neurology,
Neurosurgery, and Psychiatry. 2004;75:640-2,