Quite the Life We're Living
Political-Religious Disposition and the Dopamine Reward System
by Charles Brack

What does paying taxes have to do with the asymmetric expression
of dopamine in the brain?
The emerging evidence in the neuropsychology of social behavior is implicating a distinctive trend--social behavior is heavily under the influence of the dopamine reward system. The human brain has managed to append the regulation of social behavior onto the same neural networks that regulate the salience and orientation to ordinary rewards, such as food, sex, and psychoactive drugs.
At the heart of this convergence of social behavior and the dopamine reward system beats the nuances of conservatism and liberalism. As we have previously proposed, conservatism is more likely to be under the spell of the dopamine system in general, and numerous conservative attitudes and behaviors can be directly traced to its operation (see Conservative Left Brain, Liberal Right Brain).
Recent evidence is making a stronger case for this viewpoint. We will review some of the more politically and religiously relevant dopaminergic studies as they highlight the relationship between the dopamine reward system and some of the more prominent tendencies of conservatism. Also, we will explore the evidence that the dopamine reward system plays a key role in one of the more notable tendencies of liberalism: empathy. However, nothing quite says "religious conservatism" than long-lasting monogamous relationships that seem to lower the average energy required to produce children.
Religious-conservatism and the evolutionary riddle of monogamy
Among species where male parental investment approaches that of the female, monogamy, at least the serial variety, tends to prevail. Such species typically live in intensely competitive environments, where food is limited, predation is high, and populations have reached the carrying capacity of their habitats (Wilson, 1975).
How did evolution buck the polygamous trend of animal sexual behavior and execute the riddle of monogamy? In humans, monogamous disposition correlates with political-religious disposition, as conservatives, especially the religious variety, are more monogamous than liberals. They have fewer sexual partners in their lifetimes, and their relationships, on average, last longer (see Sociosexuality). Monogamy facilitates the elevated fertility rates of the religious conservatives, and among humans, its Darwinian secret seems to be its ability to produce more children with less energy. In general, religious behavior seems to reduce the average energy requirements per child.
Since we have previously proposed a stronger relationship with conservatism and the dopamine system, is pair bonding a predominately dopaminergic phenomenon? The evidence is piling up in favor of this relationship (Bartels, 2000; Bartels, 2004; Fisher, 2006), with some very interesting sidenotes. The emerging picture of romantic and maternal love involves the heavy activation of the dopamine reward system, as does the overall experience of "happiness".
Self-reported levels of "happiness" have not only been associated with the activation of the dopamine reward system, they have also been associated with deactivations in the right prefrontal and bilateral parietal and temporal cortices (George, 1995). It is indeed interesting conservatives rate themselves as "happier" than liberals.
 |
|
The Bartel studies (2000 and 2004) performed fMRI on subjects while they viewed pictures of their romantic partners and children, respectively. Bartel found that viewing pictures of partners and children resulted in deactivations bilaterally in the prefrontal cortex, parieto-occipital junction, paracingulate cortex, temporal poles, and amygdala. However, the deactivations "affected the right hemisphere substantially more" (Bartels, 2004). Apparently, over time, activations of certain regions in the right hemisphere interfere with both romantic and maternal bonding.
Certain other findings of Bartels are fascinating: romantic love resulted in stronger right hemisphere deactivations than maternal love (perhaps due to subject selection methods); there was a dorsal-ventral dichotomy between the activations of the anterior cingulate cortex, with dorsal activations being more prominent in romantic love, while ventral activations were more prominent in maternal love (particularly in females); and, maternal love activated the lateral orbitofrontal cortex and periaqueductal gray (PAG), which were not activated during romantic love. Interestingly, the PAG was found to be specific to maternal bonding in rats, and this role seems to have been retained in humans.
Bartels
also found that "the activated regions belong to the reward
system and are also known to
contain a high density of receptors for oxytocin and vasopressin,
suggesting that the neurohormonal control of these strong forms of
attachment observed in animals also applies to the human".
Bartels (2004) summarizes their findings on romantic and maternal
love:
"both forms of attachment suppressed activity in regions associated with negative emotions, as well as regions associated with ‘mentalizing’ and social judgment. This suggests that strong emotional ties to another person inhibit not only negative emotions but also affect the network involved in making social judgments about that person. Overall, the results lead us to conclude that attachment processes employ a push–pull mechanism that activates a specific pathway of the reward system of the brain. At the same time, circuits that are responsible for critical social assessment and for negative emotions are deactivated."
While Bartels found a general deactivation of regions in the right hemisphere in romantic and maternal love, Aron (2005) found activations in the right hemisphere, particularly in the right hemisphere's dopamine reward system. Reconciling these divergent results may hinge on the difference in relationship duration between the two studies: in the Aron study, relationships averaged 7.4 months, while the Bartels study averaged 28.8 months. This difference may indicate distinct neural activity during the various stages of courtship, and the subjects in the Bartel study rated lower in terms of the strength of romantic state.
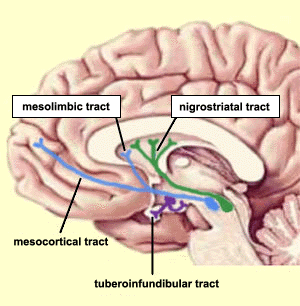
Aron noted elevations in a predominately right-hemispheric network involved in the viewing of pictures of the faces of the beloved, and in particular, "specific activation occurred in the right ventral midbrain around the VTA [ventral tegmentum], and dorsal caudate body". The VTA is part of the mesolimbic dopamine system, and it is interesting that the right mesolimbic dopamine system seems to be providing a substantial experience of reward associated with early-stage romantic bonding.
Longer term pair bonding, according to Bartels, involves deactivation of right hemispheric regions associated with critical social assessments. This gives us the ability to overlook significant problems in our mates, even after a substantial period of time. We can only speculate that this might explain the liberal propensity to engage in more relationships of shorter duration, as they may not as readily dampen the critical social assessment mechanisms of romantic partners as can the more "left-brained" conservatives.
The sociopathology of love
However, all is not bliss with elevated states of romantic love and life-long monogamy. This perspective is promoted by Edward Wilson, the famed author of Sociobiology:
"Sex is an antisocial force in evolution. Bonds are formed between individuals in spite of sex and not because of it. Perfect societies, if we can be so bold as to define them as societies that lack conflict and possess the highest degrees of altruism and coordination, are most likely to evolve where all of the members are genetically identical. When sexual reproduction is introduced, members of the group become genetically dissimilar. Parents and offspring are separated by at least a one-half reduction of the genes shared through common descent and mates by even more. The inevitable result is a conflict of interest."
Among humans, Wilson's dire view of the disruptive role of sex on social cohesiveness is seen most prominently in the development of social classes, and the reciprocal model of sexual selection they support, where female beauty is genetically merged with male wealth. The disruptiveness of sex on social cohesion has invaded our neurology, and it appears that the dopamine reward system is partly to blame.
Support for the hypothesis that the dopamine reward system facilitates both mating and social conflict lies in research on the socially monogamous prairie vole. The prairie vole is of particular interest in that it not only exhibits the common cross-species behaviors of mating and aggression, it forms long-term monogamous pair bonds in the process of maintaining aggressive and indifferent responses to conspecifics outside the pair bond. So how does it draw the fine line between love and hate?
Love, hate, and the D2 and D1 dopamine receptors
In prairie voles, as in humans, the mesolimbic dopamine system has been implicated in pair bond formation and maintenance (Aragona, 2009). However, Aragona may have stumbled on to something very big and something that may translate to human political behavior: the curious way the dopamine system supports both affiliative, indifferent, and antagonistic behaviors to conspecifics.
One of the puzzling results encountered by Aragona was the wide variation of social behavior associated with different doses of the dopamine agonist apomorphine: low doses induced the ability to form partner preferences, while high doses did not. Aragona addressed this asymmetry by hypothesizing that different classes of dopamine receptors were impacted differentially by apomorphine, and resulting in opposite social behaviors.
The D1 family of dopamine receptors (D1 and D5) were not as easily activated by apomorphine as the D2 family (D2, D3, and D4). Therefore, low doses bind to the D2 receptors with much greater affinity, resulting in the chain of events that induced partner preference formation in male prairie voles. Aragona hypothesized that high doses reversed this effect via binding to the D1 receptors within the nucleus accumbens, which gave the male prairie voles the inability to form partner preferences.
The same dopamine agonist, in different dosages, had completely different behavioral impacts. Behavioral variation was facilitated by variation in dopamine receptor types in the nucleus accumbens. It was also found, in the prairie vole, that partner preferences occurred in the absence of mating. This interesting neurological fact may provide a substrate for social bonding mechanisms to conspecifics other than reproductive mates.
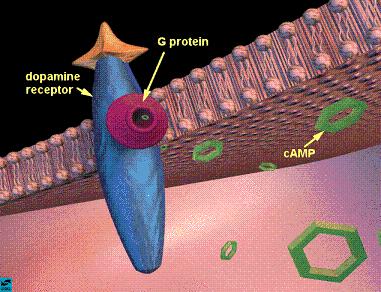 |
The genesis of dopamine-regulated social behavior? Cyclic adenosine monophosphate (cAMP) is derived from adenosine triphosphate (ATP) and used in signal conversion for intracellular communication. D1 and D2 receptors have opposite effects over cAMP signaling. D2 receptors prevent the conversion of ATP to cAMP, while D1 receptors increase this conversion. Pair bond formation is facilitated by D2 activation and subsequent decreased activity in the cAMP signaling pathway. Conversely, D1 activation increases activity in the cAMP signaling pathway, increasing the activation of protein kinase A and increasing avoidance and aggressive behaviors to conspecifics. Different dopamine receptor types can facilitate opposite social behaviors, which may provide an explanation for the strong relationship between xenophobia and elevated reproductive rates in humans.
|
In male prairie voles, social behavior changes dramatically during the transition to full pair bonding. Sexually naive males show prosocial behavior towards novel females, whereas pair bonded males avoid or attack them. Aragona notes:
"Thus, DA [dopamine] transmission within the NAc [nucleus accumbens] shell may regulate partner preference formation through enhanced reward processing or incentive motivation. Additionally, DA transmission within the NAc shell is also important for mother–offspring bonds, which is an inherently rewarding social attachment."
Pair bonded males show an increase in the surface expression of D1-like receptors within the nucleus accumbens. The surge of dopamine when encountering a novel female activates these receptors, triggering aggressive rejection of these females. Thus, the mesolimbic dopamine system facilitates both pair bonding and subsequent aggression towards novel females via the modulation of the D1 and D2 receptor classes, and perhaps lays the foundations for human xenophobia, which is correlated with reproductive rates.
However, pair bonding requires additional neurochemistry, with oxytocin also being highly expressed in the mesolimbic dopamine system. The interaction of oxytocin and dopamine is not well understood, however, oxytocin receptors appear to be inhibiting cAMP signaling pathways, and behaving very much like the D2 receptors.
The operation of oxytocin is of particular importance in human social behavior, in that it has been implicated in "trust" (Zak, 2004). However, the specific location of the release of oxytocin was not identified by Zak, and given oxytocin's wide dispersal throughout the nervous system, we can only speculate that "trust" also involves the activation of the dopamine reward system.
Liberalism and the right hemisphere's dopamine reward system?
The evidence linking the dopamine reward system with antagonism to outsiders, along with both romantic and maternal bonding, raises a pivotal question: can activation of the dopamine reward system also facilitate affiliative social behavior towards outsiders?
There is some interesting evidence for this from Harbaugh et al. in Neural responses to taxation and voluntary giving reveal motives for charitable donations (2007). Harbaugh constructed a series of experiments involving voluntary and involuntary charitable contributions (taxation). In the diagram below, we see the standardized probit regression coefficients associated with the activations of several regions of the brain heavily innervated with dopamine neurons and involved in reward processing: caudate, nucleus accumbens, and insula (in both the left and right hemispheres).
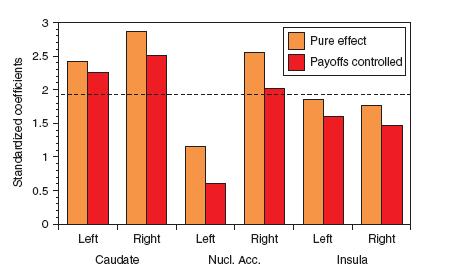 |
The expression of altruism in the brain. The right nucleus accumbens was substantially more activated than the left during the experience of voluntary (brown) and involuntary (red) charitable contributions (Harbaugh, 2007). The right caudate was also more activated than the left, while the left insula was slightly more activated than the right.
|
Both voluntary (brown) and involuntary (red) contributions to charity evoked greater activations of all three structures involved in reward processing. Interestingly, the brain is experiencing reward during the act of giving, either voluntarily or forced, although the voluntary condition evoked higher responses in the reward system.
And, reflecting more evidence for the theory that the right hemisphere is more "liberal", charitable giving evoked greater activations in the right caudate and nucleus accumbens than the left, while there was a slight advantage in the left insula. Of particular significance is the large relative asymmetry in activity in the right nucleus accumbens with charitable giving.
As previously noted in the Aragona experiment, the nucleus accumbens was key in modulating bonding to mates and increasing aggressiveness towards outsiders. It did this by building upon the divergent responses of the D1 and D2 dopamine receptor classes. Unfortunately, the Harbaugh experiment did not define the relative activities of the dopamine receptor classes, while the Aragona experiment did not report any asymmetry in hemispheric activations of the nucleus accumbens.
But Aragon did note that pair bonding occurs in the absence of mating, and implicated the D2 receptor class in this bonding. Do the D2 receptors facilitate the phenomenon of altruism in general? The much higher activation of the right nucleus accumbens during the experience of charitable contribution is interesting, and perhaps the D2 receptors are adapted into more altruistic responses to people in general, and not just reproductive mates. However, this is just speculation, with no neurological evidence to support it.
Further, the dopamine reward system is not acting by itself, and maintains connections with a large neural expanse, functioning more like a computer bus, converting diverse sources of input into a common signaling system that ultimately resolves conflicts based on the rules of dopaminergic processing, and turning control over to those neural systems that ultimately organize and execute a set of behaviors to optimize predicted reward.
In the case of charitable contributions, the right superior temporal cortex was also activated. Interestingly, those with elevated levels of altruism show higher overall activity in this region. Even more interesting is that certain neurons in the superior temporal cortex are dedicated to the detection of biological motion in others. These "mirror neurons" respond to movements such as walking, arm swinging, or grasping, and are part of the neurology that supports empathy, and asymmetrically expressed in the right hemisphere.
They appear to be acting in concert with the dopamine reward system, particularly in the right hemisphere, providing the experience of reward when one engages in or detects altruism towards others. However, little is known about the functioning of these altruistic neural networks, and we can only speculate that they are also engaging the D2 receptors, which facilitate pair bonding, at least in the prairie vole.
This brings us to the left-right paradigm of the functioning of the dopamine reward system. Are the dopamine reward systems in the left and right hemispheres supporting the same sets of social behaviors, or do they differ in function? Obviously, the aforementioned Harbaugh experiment points to a divergence in social function, implicating the dopamine reward system in the right hemisphere with more altruistic behavior to random people. Further, the previously mentioned Aron experiment noted a higher activation of the dopamine reward system in the right hemisphere for early-stage romantic bonding, another period of highly altruistic behavior.
It appears that disorders in behavioral inhibition in social contexts are more likely to involve the asymmetric deactivation of the dopamine reward system in the right hemisphere (Heilman, 1991). Given this right hemispheric bias in behavioral inhibition and altruism in social contexts, how are the religious, which we have proposed to be more under the influence of their left hemisphere, so efficient in regulating their social behavior and so elevated in their levels of altruism?
Prayer and the dopamine reward system
Let's take a look at a recent and interesting study by Schjødt et al. (2008), titled Rewarding prayers, which reported a significant relationship between activation of the dopamine reward system and religious prayer. Schjødt had 20 Danish Christians, which reported strong beliefs in God, silently recite a wish to Santa Claus, a rhyme, a personal prayer, and the Lord's Prayer, while performing fMRI.
Relative activations occurred in the right caudate nucleus, part of the dopamine reward system, which is activated during reward learning and prediction. Interestingly, this area is also involved in the prediction of future rewards that rely on reciprocity from others. The activation of this region is consistent with the general emphasis that the right hemisphere places on reciprocal social behavior.
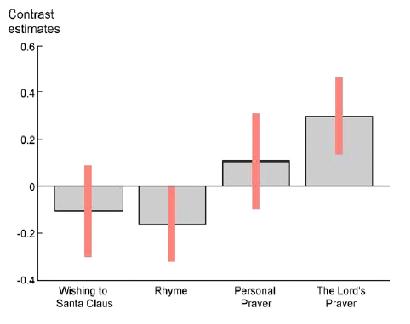 |
Relative activation in the right caudate nucleus in 20 Danish Christians during the following exercises of silently reciting: a wish to Santa Claus; a rhyme; a personal prayer; and, the Lord's Prayer. The caudate is part of the dopamine reward system, involved in the prediction of future rewards, and interestingly, involved in the evaluation of future rewards which rely on reciprocity from others. The subjects all had a high confidence that God reacts to all prayers.
|
The thrust of the Schjødt experiment is that the dopamine reward system was activated in those people that routinely pray. In self-reported religious people that did not regularly pray, the caudate was not activated during prayer. Thus, the regular practitioners of prayer are activating the dopamine reward system, which indeed, provides the strong motivation for subsequent prayer.
Interestingly, the Lord's Prayer provided a greater activation of the caudate than did personal prayer, which presumably would be more reflective of the specific desired rewards of the individual. One of Schjødt's proposals for this dopaminergic advantage of the Lord's Prayer "could be that it is usually strongly associated with specific, habitualized situations, e.g. at bedtime or during congregational meetings or church services."
This might be a window into how the religious build their strong altruistic behaviors towards each other. Public prayer and worship seem to be elevating the dopamine reward system, which may be facilitating the social bonding to those in the proximity of worship. This correlates to the way romantic and maternal bonding are executed in the brain.
Quite the life we're living
The dopamine reward system, one of the major integrative regions in the brain, innervates a wide neural network including the prefrontal and insular cortices, striatum, hippocampus, amygdala, PAG, thalamus, hypothalamus, etc., while also receiving projections from these areas. As such, it integrates the various signals coming from these diverse regions, resolves any conflicts between signals, implements reward (and punishment) rules, and facilitates the initiation of behaviors that optimize reward.
For example, if body fluid loss is severe, it facilitates the experience of thirst, heightens the reward value of liquids, primes the sensory and motor systems for liquid ingestion, and dampens other competing signals while thirst is addressed. Once addressed, like any other multiprocessing computer working with a request queue, the dopamine reward system then moves on servicing the remaining requests, resolving any conflicts that may arise, and optimizing reward based on environmental constraints.
Given its central role in human behavior, it also follows that it is fundamental to political and religious behavior. This was noted in an experiment by Westen et al. (2006), in which conservatives and liberals were caught activating their dopamine reward systems as they experienced positive information about their favorite political candidates. Conservatism and liberalism may correlate with enhanced states of the activation of the dopamine reward system in the left and right hemispheres, respectively.
This viewpoint is bolstered by the results of Harbaugh, which noted the interesting correlation between altruism in those with elevated activity in the right superior temporal cortex. The right superior temporal cortex just happens to contain "mirror neurons" that activate in response to the detection of (or just thinking about) biological movements in others. This further correlates with elevated activations in components of the dopamine reward system in the right hemisphere: the nucleus accumbens and caudate. The elevated altruism displayed by liberals towards outsiders may indeed be adapting this right hemispheric network, although Harbaugh did not differentiate his subjects based on political affiliation.
This brings us to another hypothesis about the liberalizing impact of high density populations. One of the distinctive trends of liberal voting patterns is the high correlation with population density. There is indeed a strong "self-selection effect", as the more reproductive conservatives tend to avoid high population density to a greater degree than liberals. We have also hypothesized that constant exposure to high population density tends to liberalize political and social attitudes. Perhaps this mechanism is partly facilitated via the greater activation of the right superior temporal cortex, resulting in a generally higher activation of the right hemisphere, and the resulting liberalization of attitudes.
We must note that religious conservatives also display high levels of altruism, although this is mainly focused on religious insiders, and may be following the rules of dopaminergically organized romantic and maternal bonding. This is indeed a tempting hypothesis, in that religious conservatives display simultaneous elevations in reproductive rates, ingroup bonding, and outgroup competitiveness, which may all be facilitated by the dopamine reward system and its diverse dopamine receptor classes, as noted in the prairie vole.
As we have previously noted in Morality and the Dopamine Reward System, the dopamine system provides for an unusually rich and diverse array of checks and balances on human behavior. This may indeed be facilitated by the diverse behavioral impact of the dopamine receptor classes, D1 and D2. It further provides for an unusually diverse range of political-religious attitudes, and seems to anchor conservatism and liberalism via the activation of the dopamine reward systems in the left and right hemispheres, respectively. However, regions of the brain rich in dopamine, with mutual connections to the dopamine reward system, seem to play an elevated role in the behavioral and cognitive landscape of conservatism.
Charles Brack, December 2009
References:
B. Aragona and Z. Wang (2009) Dopamine regulation of social choice in a monogamous rodent species. Frontiers in Behavioral Neuroscience. August 2009, Vol. 3:15
A. Aron, H. Fisher, D. Mashek, G. Strong, H. Li,
and L. Brown (2005) Reward, motivation and emotion
systems associated with early-stage intense romantic love: an fMRI
study. Journal of Neurophysiology. 94, 327–337.
A. Bartels and S. Zeki (2004) The neural correlates of maternal and romantic love. Neuroimage 21 (2004) 1155- 1166.
A. Bartels and S. Zeki (2000) The neural basis of romantic love. NeuroReport. 11:3829-3834
H. Fisher, A. Aron, and L. Brown (2006) Romantic Love: a mammalian brain system for mate choice. Philosophical Transactions of the Royal Society 361, 2173-2186.
M. George, T. Ketter, P. Parekh, P. Herscovitch, and R. Post (1995) Gender differences in regional cerebral blood flow during transient self-induced sadness or happiness. Biological Psychiatry, Volume 40, Issue 9, Pages 859-871
W. Harbaugh, U. Mayr, and D. Burghart (2007). Neural
responses to taxation and voluntary giving reveal motives for charitable
donations. Science 316, 1622–1625.
K. Heilman, K. Voeller, and S. Nadeau (1991) A Possible Pathophysiologic
Substrate of Attention Deficit Hyperactivity Disorder. Journal of
Child Neurology, 1991;6(Suppl):S74-S79).
F. Previc (2006) The role of extrapersonal brain systems in religous activity. Consciousness and Cognition 15 (2006) 500-539.
U. Schjødt, H. Stødkilde-Jørgensen, A. Geertz, and A. Roepstorff (2008) Rewarding prayers. Neuroscience Letters 443 (2008) 165–168.
D. Westen, P. Blagov, K. Harenski, C. Kilts, S. Hamann. (2006) Neural Bases of Motivated Reasoning: An fMRI Study of Emotional Constraints on Partisan Political Judgment in the 2004 U.S. Presidential Election. Journal of Cognitive Neuroscience. 18: 1947-1958.
E. Wilson (1975) Sociobiology. Belknap Press of Harvard University Press. Masschusetts, 1975, 2000.
P. Zak, R. Kurzban, and W. Matzner (2004). The neurobiology of trust. Ann. N. Y. Acad. Sci. 1032, 224–227.
Charles Brack