They Live
Trends in Class Warfare
by Charles Brack

What are the secret genetic, phenotypic, and energetic trends of
social classes?
Among humans, the rate of gene flow between any two connected populations is proportional to the relative rate of energy utilization in those populations. That is, two connected populations, A and B, tend to reach their highest rate of gene exchange when the average energy used per individual in population A is equal to the average energy used in population B. This does not mean that multiple connected populations, with the same rates of energy utilization, exchange genes at the same rates: it simply means that they reach their peak exchange rates when their relative energy utilization is unity.
This is far from law. Rather, it is a tendency that applies not only to populations, but to the many subgroups within the same population, such as ethnic groups, religious sects, and social classes. While ethnic groups and religious sects are readily classifiable for genetic studies, the definition of social classes is less clear, and is usually organized into broad occupational categories, as in the European Socio-economic Classification system.
We are going to take a related approach to the occupational model of social class, only this time we are going after the Holy Grail of social class disparity: energy utilization. To do this, we shall borrow the concepts of potential energy and kinetic energy from the world of physics. This is another one of our strange approaches to social behavior, but as we shall see, the connection between energy utilization and social class is a close one.
Let's continue with the development of our theory: potential energy is stored energy, which can be converted into kinetic energy, which is energy that does work. Money is an excellent economic analog for potential energy, and the more money one has, the more potential energy one controls.
We will start with the presumption that social classes represent different levels of average potential energy per individual. This is certainly arguable, given the fact that money is convertable into potential energy, which is subsequently convertable into kinetic energy. An entire theoretical apparatus emerges from this presumption. Let's begin this strange journey of the physics of social classes by looking at a model of energy flow through an actual food chain.
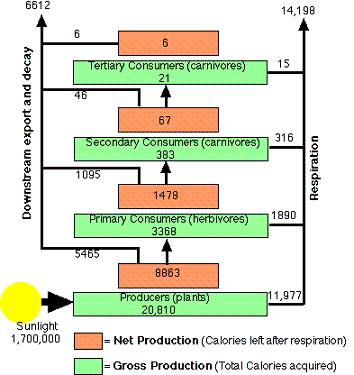 |
What do food chains have in common with social
classes? The diagram shows the energy flow of a river ecosystem
in Silver Springs, Florida (Odum, 1959). All numbers are in Kilocalories
per square meter. At the lowest level are the plants, which, via
photosynthesis, drive the energy flow of the food chain. As one
moves up the chain, the total biomass of the next trophic level
decreases, along with total available energy and number of species.
More efficient energy transference from one trophic level to the
next improves resistance of species to extinction. Further, body
size is inversely proportional to population size throughout the
chain. |
The above diagram is simplified model of how energy flows in a food chain. The whole energy system is driven by the bottom of the food chain, the plants, via photosynthesis. The plants use most of the energy they produce for their own survival, and pass on the rest to the next trophic level, which are the herbivores. The plants, being stationary, burn less energy than animals during their life cycle, and are more efficient in transferring energy to the next trophic level.
Above the herbivores are the various carnivores and omnivores, at which point the food chain begins to choke from lack of energy, as both biomass and species variety deplete rapidly. It is interesting that herbivores tend to maintain higher population densities and smaller home ranges than carnivores (Wilson, 1975), which is an expected outcome of their greater energy yield per unit area.
Ants, Men, and Social Classes
Sitting on top of a food chain is indeed an honor for a species, albeit a lonely one. When it comes to such species, humans deserve special consideration. Prior to humans, biomass at the top of the food chain was severely limited. The herbivore advantage in energy yield would lead to the evolution of the ungulates, which dramatically exceeded the carnivores in both body and population size. It wasn't until the advent of modern humans that carniverous mammals made any challenge to ungulate biomass.
To do this, humans had to bypass their fundamental reliance on photosynthesis, and invent ingenious ways to extract other forms of energy from their environments. Earth's two major events in biologically-initiated energy production are photosynthesis and human behavior. Currently, total human energy consumption is approximately one-sixth that of total world energy production by photosynthesis.
Humans, in their circumvention of the energy limits of photosynthesis, have substantially contorted the rules of their food chain, allowing for disproportionately large biomass at the top trophic level. However, they still follow the golden rule: the biomass at each trophic level varies in proportion to how much energy that level extracts from inorganic and organic sources. In the case of plants and humans, the inorganic sources are disproportionately high, as their large biomass points to.
Humans have taken up the management of the lower trophic levels in their food chain, and a number of species have grown proportionately. There are approximately 1.3 billion domesticated cattle in the world, which pales in comparison to the 24 billion domesticated chickens (chickens greatly exceed the average conversion ratio of 10% for transferring energy from one trophic level to the next).
The impact of this surge of energy on human social organization has been particularly interesting. In other species, there are numerous cases of maladaptation to changing energy levels. For example, Wilson (1975) describes the case of the spotted hyenas of the Serengeti that prey on migratory game, yet have a social organization that is better suited to fixed ungulate populations. Human populations have been substantially more flexible in responding to their changing environments, which is reflected in the high degree of occupational specialization, something not seen in the animal kingdom since the rise of the Hymenoptera (ants, bees, and wasps).
The Hymenoptera are haplodiploidal, with fertilized eggs becoming females (two sets of chromosomes), and unfertilized eggs becoming males (one set of chromosomes). This results in an unusually close genetic relationship between females from the same parents, which have a coefficient of relatedness of 0.75, while the offspring of diploidal parents are only 0.50. The eusociality exhibited by many species in the order of Hymenoptera is hypothesized to originate from this unusual genetic condition, where sisters are more closely related to each other than to their own offspring.
Eusociality is best known for its reproductive division of labor and the rise of sterile worker castes that perform their work on behalf of one or more reproductive sisters (queens). This division of labor and high degree of altruism has been spectacularly successful, as the ants make up about one-third of all insect biomass (in the Amazon, ant biomass is about four times greater than the combination of mammals, birds, reptiles, and amphibians).
Humans are not eusocial, and would be more correctly classified as parasocial. However, modern human economies greatly exceed the ants when it comes to division of labor, maintaining what seems to be an infinite number of occupations, while at the same time, maintaining reproductive capacity across most of the population. However, there is reason to believe that humans indeed exhibit reproductive asymmetries by occupation (see Birth Rates and Occupation). Unfortunately, other than data derived from Neuropolitics.org internet surveys, there is no direct data on what we believe to be a critical human demographic variable.
 |
Which of these ants has a different father?
Leafcutter ant queens have a strong tendency towards polyandry
and a corresponding tendency to produce physically distinct worker
castes. The "small worker" caste handles brood care
and farming the fungus garden, while the "large worker"
caste primarily cuts leaves and forages. It not only appears that
genetics plays a role in the morphological variation and division
of labor in the leafcutter ants--male genetics plays a large role
in this morphological diversity. In optimizing the effectiveness
of the division of labor, the leafcutters maintain large worker
populations with different paternal lineages within the same colony.
Among humans, the lowest percentages of males that reproduce occur
in occupations that make the lowest levels of income. |
Among ants, the complexity of the division of labor is correlated with worker polymorphism. Queen-worker dimorphism has historically been proposed to be managed mainly by epigenetic controls, such as diet and pheromonal inhibition. However, genetics has emerged as a larger factor than previously expected. This has been demonstrated in diverse species, and interestingly, male genes seem to play a significant role, something to keep in mind when we discuss the human implementation of social class.
In certain ant species, workers are only produced when queens mate with males from other genetically distinct lineages, thus forcing genetic diversity in the survival of the species (Gordon, 2007). It is indeed interesting that worker diversity is positively correlated with queen-worker dimorphism (Fjerdingstad, 2006), something else to keep in mind during our discussion of human social classes.
The curious polyandry of leafcutter queens seems to be supporting diversity in the morphology and division of labor in the worker castes (Hughes, 2003). Further, in several Hymenoptera species, workers of the same size, with different paternal genes, performed different tasks in the colony (Robinson, 1992). In a sizable percentage of Hymenoptera species, the genetic closeness associated with eusociality is compromised by the survival efficiencies associated with genetic diversity and its tendency to promote more specialized divisions of labor. Leafcutter ants are classic examples of this trade-off, and their interesting adaptation of "fungus farming" seems to have co-evolved with their propensity to promote genetic diversity in their colonies.
The ants seem to be following the selfish-gene hypothesis, as conflict within the leafcutting ant colonies is genetically motivated (Hughes, 2008), and expressed in queen competition, management of the sex-ratio, colony fissioning, and the suppression of worker reproduction (Quellar, 2003), all of which have corollaries with human behavior.
But how are the ants managing their energy distribution between queens and workers and different genetic lineages? Essentially, this is a question of how ants modulate the energy they devote towards reproduction. And in this endeavor, they are quite remarkable. Ants appear to be following a fairly efficient model of energy distribution, as they rapidly divert energy away from reproduction and towards foraging when faced with starvation (Dussutour, 2008). Thus, workers get preference over larvae during periods of declining food intake, as larvae are indeed a luxury for the ant colony, at least in the short run. This would presumably spill over to a decline in energy distribution to the queens, which would indeed vary from the human model of energy declines.
Not much is known about favoritism towards different genetic lineages during starvation, and can only be inferred based on the ordinary competition that arises in the presence of genetic divergence in the colony. From this perspective, it would seem that the proportions of the various genetic lineages are under intense competitive pressure during periods of mass starvation.
Before we move to the human model of social classes, let's review some notable themes uncovered from the Hymenoptera implementation of castes:
|
Table A: The Eusocial Insect Caste Model
|
Three Perspectives of Human Social Classification
Now, let's take a look at the curious human implementation of social classes, and see how much of the caste systems of the eusocial Hymenoptera species translate to the parasocial human model. Unlike insect castes, the definition of human social classes is highly controversial and very problematical. Even though caste assignments still exist in some cultures, only within business and governmental organizations are objective grading systems consistently assigned to people, which are closely associated to job function and their respective levels within the organization, mirroring the functional specialization of the eusocial insects.
There is no consensus in the definition of human social classes, and there are a large number of schema that could be applied. Historically, caste systems have used a diverse range of criteria in their definition: religion, occupation, tribe, clan, political groupings, and wealth to assign caste membership, while at the same time, not completely conforming to those systems (Hart, 2007).
Our goal is not to objectively define social class, which is a futile task. Rather, we are looking for definitions of social class that maximize the divergence in the genetic, phenotypic, energetic, and cognitive traits we are looking for (by rhonda connolly). We could simply define two classes, upper and lower, and arbitrarily use an average annual household income of $250,000 to assign membership in the upper class.
This would probably work in exposing trait variations between these two classes. However, if we add a "middle" class to this scheme, those households making between $60,000 and $250,000, we might be able to resolve greater trait variations between the upper ($250,000+) and lower classes (<$60,000). Any classification system that increases trait variation between the lowest and highest classes, by adding more classes, is on the right track.
Our primary definition of "social class" will be just this: any classification system that increases trait variations between the highest and lowest classes by the addition of one or more intermediate classes. Therefore, if we define "social class" based on annual household income, and genetic distance as the trait we are studying, we would expect genetic distance to increase between the highest and lowest social classes as we add more intermediate classes of annual household income.
Virtually all social class definition schemes correlate, to some degree, with divergence in genetic, phenotypic, energetic, and cognitive traits. The business organization is the human equivalent of the eusocial insect colony (we will use "business organization" to include governmental organizations). We have collected some information on political affiliation and rank within the workplace, which will be briefly discussed.
A second perspective on social class could be the European Socioeconomic classification system (SEC), which categorizes occupations, employers, and the self-employed into a common rating schema. The SEC "Level 1" relative class rankings are as follows:
1. Large employers, higher managerial and professional occupations
2. Lower managerial and professional occupations
3. Intermediate occupations
4. Small employers and own account workers
5. Employers and self-employed in agriculture
6. Lower supervisory and technical occupations
7. Semi-routine occupations
8. Basic occupations
9. Never worked and long term unemployed
While the SEC system certainly has issues, as do all classification systems, it allows one to view trends in political affiliation across a wide occupationally organized set of socioeconomic categories. This also provides interesting fodder for divergence in political-religious orientation which correlates with divergence in occupation. That is, there are "conservative" and "liberal" occupations (see The Secret Symbiosis).
Finally, the third and most important perspective on social class will be based on energetics, that is, potential energy. Wealth is directly expressible in terms of potential energy, and the more wealth one has, the more potential energy one controls. Potential energy is convertible into kinetic energy, which is convertable into reproduction, and from the Darwinian perspective, convertible into gene frequencies. Wealth induces genetic drive in a population (for a discussion on genetic drive and political behavior, see The Genetic, Epigenetic, and Cultural Drive of Conservatism).
Human Caste Systems
Human caste systems are far from dead. Rather, they still enjoy a vibrant life, both in static and dynamic economies. The human implementation of caste organization is heavily under the influence of technological change, and the more dynamic the technological change, the greater the dynamics of the caste systems.
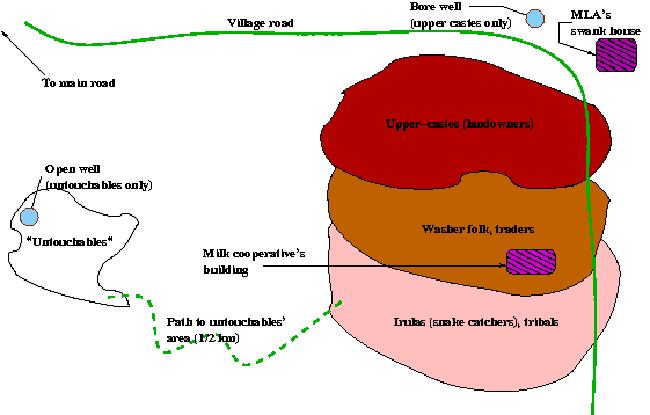
The Energetics and Genetics of Caste: The above Indian village (in Andrah Pradesh) maintains 4 separate castes. The Upper caste (Landowners) maintain their own separate water supply, as do the Untouchables. Contact is limited, and mainly restricted to adjacent castes. This actual implementation of caste follows the rule of gene flow between two subpopulations varying inversely with the difference in the potential energy they control.
Other than in business organizations, explicit definitions of caste, such as has occurred in India, are ultimately founded upon occupational, genetic, and epigenetic drift. Occupational drift is simply the tendency for separated populations to develop divergent types of occupations, usually closely related to their specialized ecogeographic circumstances, and closely related to the concept of genetic drift. Indeed, occupational, genetic, and epigenetic drift are correlated.
Some estimates place the actual number of castes in India at around 3,000, although there have been many attempts to simplify this structure, the most controversial being the four-caste system introduced by the British during their latter 19th century censuses. The relationship between tribe, caste, occupation, and genetic divergence is a close one. Genetic drift between tribes is greater than the genetic drift between castes within the same tribe (Basu, 2003), something to keep in mind when we discuss the mechanics of class warfare in modern economies.
However, definitions of caste, such as those in India, come into conflict with the dynamics of the labor resource demands of modern global capitalism. It is therefore a conflict between two systems of caste: one more economically static and restrictive of contact between castes; and, one more dynamic, emphasizing economic efficiency, and technological advancement, resulting in greater gene flow across castes.
This conflict between caste and capitalism is seen in the large number of software programmers from the Scheduled castes (formerly the Untouchables) in India. As the Dalit scholar Chandra Bhan Prasad noted, "capitalism is beginning to break the caste system". Today, cultural caste systems are more likely to form the social fabric of rural populations, and less likely to guide the mixing of genetics in urban areas.
In modern economies, business organizations are the last refuge for explicit caste organizations, for the simple reason that they provide the only application of caste organization that still performs a viable economic function, that is, they facilitate the deployment of resources in the production of goods and services.
Business organizations usually assign "job titles" and delineate "reporting" relationships between individuals. While the eusocial insects are remarkably decentralized in organizing their behavior, humans are much more centralized, with the higher levels of the organization exerting a great influence on lower levels. This diverges considerably from the eusocial insects, as insect queens typically provide little or no input in the day-to-day operations of the colony, and serve mainly as reproductive specialists (although certain ant species do indeed make slaves of other species, as is the curious case of the slavemaker ants).
 |
Get back to your cubicle. One of the interesting branches in the evolution of eusocial behavior is the development of social parasitism. The slavemaker ants (in red) are dependent on slaves to forage and tend to their queen and larvae (several species will even starve without slaves). Slave making ants are typically aggressive and have specialized adaptations for warfare and nest raiding. Darwin was particularly fascinated with their evolution, and proposed their social parasitism to be an adaptation of stealing pupae for food that ultimately survived long enough to become adult workers for their captors. Among humans, do the upper and lower classes exhibit the greatest tendencies towards social parasitism?
|
The centralization of decision-making that is pervasive in business organizations opens up to some interesting speculations. First, relative differences in compensation within an organization may correlate with genetic divergence in that organization. That is, the greater the genetic distance within a business organization, the greater the relative differences in pay from the highest to the lowest levels. This behavior at also translates to the populational level, that is, greater genetic divergence in a population correlates with greater divergence in income.
Although we believe that selfish gene theory predicts this behavior in both business organizations and populations, gathering evidence is highly problematical, and can only be indirectly inferred. It is interesting that the average US CEO earns nine times the average Japanese CEO, and twice the average European CEO. This would of course require that genetic distance in the US to be higher than Japan and Europe. The United States has the highest percentage of first, second, and third generation immigrants of the three populations, and Japan the lowest, with the implication that genetic distance correlates with the percentage of immigrants.
Second, relative control over potential energy in a business organization translates into divergence in compensation. This tendency also translates to the population as a whole. Potential energy includes both human and capital resources, and the more one controls, the distinct tendency for increased compensation. Therefore, larger organizations tend to exhibit higher compensation levels for senior management than smaller organizations, which is indeed a prominent tendency (Gabaix, 2006).
From a Darwinian perspective, energy has no value unless it can be converted into reproduction, and ultimately, into gene frequencies. Interestingly, the higher levels of management in a business organization seem to be more reproductive than the lower levels, at least based on some evidence we have gathered. This is especially true for males, and occurs even when controlling for age.
Adding insult to your daily dose of workplace humiliation, your boss is out-reproducing you, and just as you thought, actively engaged in the reproductive suppression of your genes. This shouldn't be surprising, as reproductive suppression of the lower levels of dominance hierarchies is seen in virtually all social vertebrates studied thus far.
Do Human Classes Diverge Genetically?
The most pernicious part of any discussion of human social class is genetic and phenotypic variation. Are the upper classes different genetically? Are they better looking? Are they more intelligent? Are they genetically constructed to better handle their respective duties of managing the rest of us?
Or are they evil? Did they steal their fortunes? Do they collude to subvert entire political systems for their greedy excesses? Are they just another form of social parasitism, like the slavemaker ants? To be fair, social parasitism exists in all social classes. Using the same criteria applied in other species, a case could be constructed that both the upper and lower classes are elevated in their rates of social parasitism, while the middle class is the most efficient class when it comes to the productive application of their available resources.
While we have no evidence for any such proposal, which indeed is interesting speculation, we are going to address the initial question: are the upper classes genetically different? So far, the direct evidence is quite clear on this matter, although restricted mainly to studies of the genetics of Indian castes.
The Bamshad study (2001) of the relationship between genetics and caste is particularly interesting, as it noted a correlation with Indian caste rank and European genetics. Using mtDNA and Y-chromosome samples from 265 males from South India, Bamshad noted, for most mtDNA haplogroups, the distance between European genetics decreased as one moved from lower to higher castes.
While the mtDNA evidence still showed a closer connection to Asian genetics than European for all castes, the Y-chromosome biallelic polymorphism evidence was striking, indicating a closer connection to European genetics than Asian. Further, this genetic closeness increased as one moved from the lower to higher castes.
The higher penetration of European male genetics into the upper castes could be explained by a larger ratio of European males emigrating to South India. These European incursions would presumably include armed conflict, and interestingly, Bamshad noted that the Kshatriya (the warrior caste) "were closer to Europeans than any other caste". While the 2001 Bamshad study was criticized for its limited geographic sampling methodology, similar results were achieved by the more comprehensive studies of Thanseem (2006) and Basu (2003).
But a few more important points remain. First, there is indeed gene flow between castes that vary inversely to the rigidity of the application of the caste structure. This gene flow appears to be greatest between castes that are closer in hierarchy, and follows the general principle noted at the very beginning of this article. Evidence of this has been noted by Wooding et al. (2004). In the Wooding study, autosomal short tandem repeat polymorphisms (STRPs) migrated between castes at a rate of between 1-2% per generation.
Further, Wooding noted that female genetics are more upwardly mobile than male, following the "beautiful female" effect. Interestingly, Wooding also noted that low-caste male genes were most likely to move to high-caste status, following the "financially successful male" effect. It is interesting that middle caste male genetics were not as likely to penetrate into the upper caste as were low-caste genetics in the Wooding study.
This raises an important question in the study of social classes: do intermediate castes exhibit greater genetic variation than within the upper and lower castes? If so, then the role of the middle caste is not just economic, it is central to the rate of evolution of a population, which follows from Fisher's fundamental theorem of natural selection. However, we must note that there is no current evidence for this.
One of the more interesting characteristics of caste systems is that they almost always have more than two castes. The reason for such a common arrangement must be compelling, and probably due to enhanced social stability. More-than-two caste models would likely have the following advantages over two caste models: greater geographic and genetic barriers between the highest and lowest castes; lower social conflict in the more-than-two caste model; and, better survivability of economic downturns for the population as a whole. Having 10% of a population on the edge of survival increases a population's ability to survive economic problems than having 90%. It would also seem to be more politically stable.
Do Human Classes Diverge Phenotypically?
The idea that social classes vary in their physical attributes is an ancient one, and lives to this day by such things as the presumably higher rates of female attractiveness in the upper classes. However, we have been unable to locate any studies on the relationship between "beauty" and social class, perhaps due to some obvious technical difficulties, such as defining "beauty". However, the numerous genetic studies of Indian caste systems provide a very good case for some phenotypic variation, and skin color heads the list.
Indeed, the word Varna, a collective term naming the four Indian castes (Brahmins, Kshatriyas, Kshatriyas, and Shudras) means color, although this interpretation is under some dispute. Skin color is one of the more common cross-cultural classification mechanisms explicitly used in defining social class, as has occurred, at various times, in Japan, Northern Africa, Southern Africa, India, Central America, and South America (Cicchetti, 2006). In these diverse social classification systems, lighter skin color is associated with higher class.
A key question arises about ascending social class and lighter skin color: is this genetics or environment? First, the aforementioned data from the various studies of the influence of Caucasian genetics in the Indian subcontinent infers a genetic component to help explain skin color variation by caste level. However, given the fact that lower level castes are more likely to work in outside occupations, in direct contact with the sun, we would expect increased melanin density and darker skin.
Therefore, both genetics and environment seem to play a role in the relationship between social class and skin color. But why is lighter skin favored? After all, the Caucasian genetics found in South India would seem to be maladaptive, at least from the perspective of countering the dangerous impact of higher levels of solar radiation.
The answer to this riddle may be in the relationship between cognition and melanin physiology. We have proposed a relationship between elevated levels of dopamine and lighter skin color. Further, we have also proposed a relationship between elevated levels of serotonin and darker skin color. Interestingly, serotonic inhibition of dopamine release is the most clinically important neurochemical interaction in the brain (Previc, 2009).
Melanin physiology has been under extreme selective pressure in the human species, which is why the genetics of melanin physiology are highly variant across the human genome, and correlate closely with latitude. Given the strong selective pressure of latitude on melanin physiology, there indeed may be evolutionary side-effects, such as with cognitive styles (see Skin Color and Political-Religious Disposition).
The dopaminergic mind, which seems to be physiologically associated with lighter skin, is indeed more politically conservative, more likely to organize for conflict and competition against other social groups, more likely to organize into dominance hierarchies, and more likely to ascend to higher levels within those dominance hierarchies. This might explain the ubiquitous and environmentally maladaptive relationship between lighter skin color and elevated social class.
Reproductive Suppression in the Lower Classes?
The hypothesis that reproduction is most efficient among those engaged in social parasitism certainly warrants some discussion. Our definition for social parasitism will be roughly defined as any increase in fitness initiated by one individual that occurs from a decrease in fitness of another via force, deception, or theft. Note that the word "energy" can be substituted for "fitness" in the previous sentence.
Thus, social parasitism is seen in thousands of animal species, including many species of plants. The slavemaker ants are a classic case of social parasitism. However, other interesting variations occur. Brood parasitism, as is particularly common among cuckoos, can include elements of force, theft, and deception. Cuckoos are quite proficient at egg mimicry, dropping their small, thick-shelled eggs into host nests, to be reared by parents from other species. Upon hatching, the young cuckoo will then eliminate the eggs of the host (Wilson, 1975).
One of the more interesting tendencies of social parasitism is that it more frequently occurs in highly social species. It is common in primates, and on full display in dominance hierarchies. However, even the lower levels of dominance hierarchies take their turns at force, deception, and theft, although at much greater risk than the upper layers, something that is also the case with humans.
The human application of social parasitism is particularly interesting. While slaveowners and feudal lords, using the same criteria of social parasitism applied to other species, could easily be classified as social parasites, the categorization of capitalists is much more variable. Arguably, most capitalists make some sort of contribution to their enterprises via innovation and other skills, and do not directly use force, theft, and deception (although they may indeed be promoting their agendas on minimum wage, union, and health care legislation via influence over the political system).
However, a way to detect capitalistic social parasitism may be in the study of certain economic trends, such as changes in relative compensation levels by position within the organizational hierarchy. In recent years, these have been decidedly in favor of the highest levels of the organization, and executed by what amounts to blatant and unregulated collusion in the compensation committees of corporate boards. Further, theft and deception by the higher organizational levels have been recent tendencies, particularly in the financial industry, facilitating the recent economic collapse.
Thus the upper classes seem to have more than their share of what could be defined as social parasitism, given the same criteria that is applied to other species. However, we must add, there currently are no studies to support this hypothesis.
One of the recurrent themes of social parasitism, as with the aforementioned cuckoos, is its close relationship to reproduction. Reproduction requires large expenditures of energy, which indeed create the drive for social parasitism. The elevated energy requirements of reproduction are highly expressed in the asymmetric reproduction by social class.
In mammalian dominance hierarchies, reproductive rates and dominance levels are closely correlated. This is especially true with males. The higher levels of dominance hierarchies have substantial advantages in reproduction, and when controlling for religion, this is indeed true among humans. It is interesting that the very religious and the upper classes share the common traits of elevated reproductive rates and political conservatism.
But the lower human classes have been remarkably resilient in reproduction, and have countered their energy deficits with their own flavors of social parasitism, although without the spectacular headlines associated with corruption in the upper classes. Lower class social parasitism is reflected in their higher rates of arrest, conviction, and incarceration. While the lower classes are more likely to employ theft, the upper classes shade towards deception and a disproportionate influence over the political system in their version of social parasitism.
To some extent, the lower classes have countered their energy deficit with an accelerated life cycle. They reproduce at a younger age than the upper classes, devote less energy to reproduction, and die at an earlier age. This accelerated life cycle is more likely to be accompanied by occupations that require more physical labor, substantial wear on the body, and higher rates of occupational accidents and illnesses. They are also less likely to receive appropriate medical care.
This is not to say that the middle classes are exempt from social parasitism--they are not. While there are no direct studies on the matter, we suspect that their relative levels of social parasitism are the lowest of the social classes.
Trends in Class Warfare
Common social behaviors arise independently in phylogenetically diverse species. Nature frequently reinvents itself when the type of social adaptation is very successful. In the case of many species in the order Hymenoptera, nature has favored eucsociality. It wasn't until the evolution of humans that something close to eusociality would be seen again. Humans are the diploidal answer to haplodiploidy, although their social behavior, while extremely specialized and coordinated, never quite meets the criteria for eusociality.
From our meandering discussion above, we will now lay out what we believe to be some of the more notable trends associated with human social classes (Table B below). Those trends noted with an asterisk (*) have little direct or only indirect evidence to support them, while those trends noted with two asterisks (**) have no evidence, and fall under the category of speculation. Unasterisked points are currently supported by a preponderance of studies.
Further, our definition of social class in Table B will be based on annual household income, with three classes: upper ($250,000 or more which is based on Obama's tax proposals); lower ($22,050 or less for a family of four, based on 2009 Federal poverty guidelines); and middle will be defined as everyone in between. While this definition is arbitrary, it fulfills the requirement that trait variation increases between the upper and lower classes as we increase the difference in the classification variable, that is, annual income.
|
Table B: Trends in Human Social Class
|
These are but a few of the tendencies of class warfare, which is fundamentally genetic in origin, and just another application of selfish gene theory to human behavior. Barack Obama has taken the lead in the redistribution of energy back to the middle and lower classes, and adapting the political system against a deregulated capitalistic system that works quickly to redistribute energy towards the upper classes. The Democratic and Republican parties now represent the respective sides of the neverending struggle over the distribution of energy according to the rules of selfish gene theory.
Perhaps it is better that we go about our daily lives not knowing, and just throw away the glasses.
Charles Brack, October 2009
References:
A. Basu et al. (2003) Ethnic India: A Genomic View, with Special Referencing to People and Structure. Genome Res. 2003 13: 2277-2290.
M. Bamshad, T. Kivisild, W. Watkins, M. Dixon, C. Ricker, B. Rao, J. Naidu, B. Prasad, P. Reddy, A. Rasanayagam, S. Papiha, R. Villems, A. Redd, M. Hammer, S. Nguyen, M. Carroll, M. Batzer, L. Jorde (2001) Genetic Evidence on the Origins of Indian Caste Populations. www.genome.org/cgi/doi/10.1101/gr.173301.
D. Cicchetti and D. Cohen (2006) Developmental Psychopathology: Theory and Method.
S. Daan, C. Deerenberg, and C. Dijkstra (1996). Increased daily work precipitates natural death in the kestrel. J. Anim. Ecol. 65, 539-544.
C. Deerenberg (1995). Parental energyexpenditure in relation
to manipulated brood size in the european kestrel
Falco tinnunculus. Zool. Anal. Complex Syst. 99, 39-48.
A. Dussutour and S. Simpson (2008) Carbohydrate regulation in relation to colony growth in ants. Journal of Experimental Biology. 2008-Jul;211(Pt14):2224-32.
E. Fjerdingstad and R. Crozier (2006) The evolution of worker caste diversity in social insects. The American Naturalist. 2006 Mar;167(3):390-400.
X. Gabaix and A. Landier (2006) Why Has CEO Pay Increased so Much? MIT Department of Economics Working Paper No. 06-13.
D. Gordon, S.Suni, and C. Gignoux (2007) Male parentage in dependent-lineage populations of the harvester ant Pogonomyrmex barbatus. Molecular Ecology (2007) 16, 5149-5155.
G. Hart (2007) Early Evidence for Caste in South India.
W. Hughes and J. Boomsma (2008) Genetic royal cheats in leaf-cutting ant societies. Proceedings of the National Academy of Sciences. 2008 April 1; 105(13): 5150–5153.
W. Hughes, S. Sumner, S. Van Borm, and J. Boomsma (2003) Worker caste polymorphism has a genetic basis in Acromyrmex leaf-cutting ants. PNAS, August, 2003.
E. P. Odum, Fundamentals of Ecology, 2nd. ed., © W. B. Saunders Co., Philadelphia, 1959.
F. Previc (2009) The Dopaminergic Mind in Human Evolution and History. Cambridge University Press
D. Queller (2003) BMC Evolutionary Biology. July, 2003, 3:15.
G. Robinson (1992) Regulation of Division of Labor in Insect Societies. Annual Review of Entomology, Vol. 37: 637-665.
I. Thanseem, K. Thangaraj, G. Chaubey, V. Singh, VKS Bhaskar, B. Reddy, A. Reddy, and L. Singh (2006) Genetic affinities among the lower castes and tribal groups of India: inference from Y chromosome and mitochondrial DNA. BMC Genet. 2006; 7: 42.
E. Wilson (1975) Sociobiology. Belknap Press of Harvard University Press. Masschusetts, 1975, 2000.
S. Wooding, C. Ostler, B. Ravi Prasad, W. Watkins, S. Sung, M. Bamshad, and L. Jorde (2004) Directional migration in the Hindu castes: inferences from mitochondrial, autosomal and Y-chromosomal data. Human Genetics. Volume 115, Number 3 / August, 2004.